INTRODUCTION
Marine algae are valuable source materials for about 2,400 natural products with pharmacological and biological activities (Faulkner 2001, Fatima et al. 2016). These potential source materials are used as antibiotic, antitumor, antioxidant, antifouling, and antiulcer products, as well as suspension agents in radiological preparations (Athukorala et al. 2006, Oh et al. 2016, Park et al. 2016, Fernando et al. 2017). For example, the red alga Portieria hornemannii (Lyngbye) P. C. Silva has been evaluated for its biological (e.g., antibacterial and antioxidant) activities. (Fuller et al. 1992, 1994, Fatima et al. 2016). Its pharmacological activities are usually attributed to two secondary metabolites, apakaochtodenes A and B (Puglisi and Paul 1997), and halogenated monoterpenes, which are effective at deterring herbivores (Paul et al. 1990, 1992). According to studies of variation in the chemical composition of secondary metabolites, marine algae produce diverse mixtures of secondary metabolites among sibling species (Masuda et al. 1997, Vairappan et al. 2014). Moreover, the production of secondary metabolites notably differs within the same species found at different sites (Matlock et al. 1999, Oliveira et al. 2013). For instance, extracts of P. hornemannii collected from six different sites on Guam contained significantly different concentrations of major secondary metabolites (i.e., apakaochtodenes) (Puglisi and Paul 1997), possibly due to site-specific environmental influences. Considering that the synthesis of different secondary metabolites occurs via different metabolic pathways, the compositional and spatial variations in secondary metabolites in P. hornemannii imply variable genetic diversity masked by morphological similarities (Masuda et al. 1997).
Portieria is a genus in the red algal family Rhizophyllidaceae in the order Gigartinales, characterized by thalli up to 15 cm long that are flattened, irregularly bipinnate or tripinnate, and typically bushy (Payo et al. 2013). Of this genus, two species, P. hornemannii and Portieria japonica (Harvey) P. C. Silva, have been reported in the northwest Pacific (Okamura 1936, Kang 1966, Silva et al. 1987, Yoshida 1998, Lee and Kang 2002). P. hornemannii, a currently accepted type species in the genus Portieria, is thought to have a broad distribution in Africa, Australia, the Pacific Islands, and Asia (Silva et al. 1987, Yoshida 1998, Lee and Kang 2002, Guiry and Guiry 2018). Although recent morphological information including reproductive development have been supplemented earlier descriptions, the morphology of most Portieria species remains to be characterized in detail (Payo et al. 2011). P. japonica, initially reported as Desmia japonica Harvey collected on rocks at low tide from Shimoda, Japan (Harvey 1860), has been demonstrated to have narrower distributions in the western Pacific, including Korea, Japan, China, Taiwan, the Philippines, and Vietnam, as well as Oman in the Middle East (Silva et al. 1987, Yoshida 1998, Lee 2008, Guiry and Guiry 2018). Morphologically, P. hornemannii has branches that gradually narrow toward the apices, whereas P. japonica is distinguished by relatively broad branches with sawtooth margins (Yoshida 1998, Payo et al. 2011). However, it is relatively difficult to discriminate between P. hornemannii and P. japonica based on morphological characterization alone (see Chondrococcus hornemannii and Chondrococcus japonica in Okamura 1922).
Recent molecular methods have clarified numerous taxonomic questions arising from morphological similarities; this has resulted in the discovery of a vast number of previously unaccounted hidden species (Tronholm et al. 2010, Payo et al. 2013, Lee et al. 2016). For example, a recent molecular study of P. hornemannii confirmed that the current assessment of its species diversity in the Philippines archipelago was considerably underestimated (Payo et al. 2013). At least 21 cryptic species of Portieria were found to be geographically structured in areas smaller than even one archipelago (<100 km), reflecting the possibility of speciation at a fine spatial scale (Payo et al. 2013). Moreover, it reveals the potential for hidden diversity of Portieria in other regions, particularly the northwest Pacific, where is the place to detect genetic diversity of P. japonica besides those of P. hornemannii.
In this study, we investigated the genetic diversity of Portieria in Korea, Japan, and Taiwan. To verify the existence of cryptic species, we analyzed sequences of the plastid rbcL gene and evaluated the genotype distribution of Portieria species in this region. We also inferred phylogenetic relationships of the members of Portieria.
MATERIALS AND METHODS
Portieria specimens were collected from the low intertidal to subtidal zones at 14 sites in Jeju, Korea, 4 sites in Japan, and 1 site in Taiwan (Table 1). The specimens were initially kept in a plastic box cooled with ice and transported to the laboratory. The materials were pressed onto herbarium sheets, whereas those materials used for molecular studies were desiccated in silica gel. The locality information and GenBank accession numbers are listed in Table 1. Photomicrographs of the general plant habit were captured using an EOS 600 D digital camera (Canon, Tokyo, Japan). Voucher specimens were deposited in the herbarium of Jeju National University (JNUB, Jeju, Korea).
Genomic DNA was extracted from the dried thalli ground in liquid nitrogen using a DNeasy Plant Mini kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. The plastid rbcL gene was amplified by polymerase chain reaction (PCR) using AccuPower PCR Premix (Bioneer, Daejeon, Korea). The PCR protocol for rbcL amplification included initialization at 96°C for 4 min followed by 35 cycles of denaturation at 94°C for 1 min, annealing at 50°C for 1 min, and extension at 72°C for 2 min, with a final extension at 72°C for 7 min. The rbcL primers used for amplification and sequencing were as follows: F145, F762, R898, and R1381 (Freshwater and Rueness 1994, Kim et al. 2010). The PCR products were purified using an AccuPrep PCR Purification Kit (Bioneer) according to the manufacturer’s instructions. Sequencing of the purified PCR products was performed by Macrogen (Seoul, Korea).
All sequences were determined in both forward and reverse strands, and the electropherograms were checked and edited manually using Chromas ver. 1.45 (Queensland, Australia). Nucleotide sequences were aligned using BioEdit (Hall 1999). To construct the phylogenetic tree, the sequences were aligned with those of related species downloaded from GenBank: P. hornemannii from South Africa (U26825, U04215), P. japonica from Japan (U26825, U04215), and Portieria tripinnata (Hering) De Clerk from South Africa (EU349205). To compare partial rbcL sequences of Portieria species from the Philippines, we aligned 355 bp of the rbcL gene between our specimens and 21 representative haplotypes in Payo et al. (2013). Ochtodes secundiramea (Montagne) M. Howe (KJ404066) and Contarinia peyssonneliaeformis Zanarnidi (EU349200) from the family Rhizophyllidaceae were selected as outgroup species (Mendoza-González et al. 2011). To infer the phylogenetic relationship, we performed a maximum likelihood analysis using the GTRGAMMA evolutionary model in RAxML ver. 7.2.8 (Stamatakis 2006). To identify the best-fitting tree, we performed 200 independent tree searches using ‘the number of runs’ option. We ran 1,000 bootstrap replications in RAxML using the same setting to generate the support value of monophyletic nodes (MLBt). We also performed Bayesian inference in MrBayes ver. 3.2.1 (Ronquist et al. 2012) using metropolis-coupled Markov Chain Monte Carlo simulations with the GTRGAMMA model to select the most appropriate phylogenetic tree. For two independent runs, we computed 2 × 106 generations with four chains, and sampled the trees every 100 generations. The burn-in point was identified graphically by tracking the likelihoods when they plateaued. Eventually, 19,750 trees sampled in the stationary state were used to infer the Bayesian posterior probabilities (BPP). Gene genealogies were estimated with a statistical parsimony network in TCS 1.21 (Clement et al. 2000) and a minimum spanning network in ARLEQUIN ver. 3.5.2 (Excoffier et al. 2005).
RESULTS
In total, 76 rbcL sequences (1,228 bp), including 28 sequences obtained from GenBank, were used for the alignment. Variable and parsimoniously informative sites were found at 420 (34.2%) and 263 (21.4%) positions, respectively.
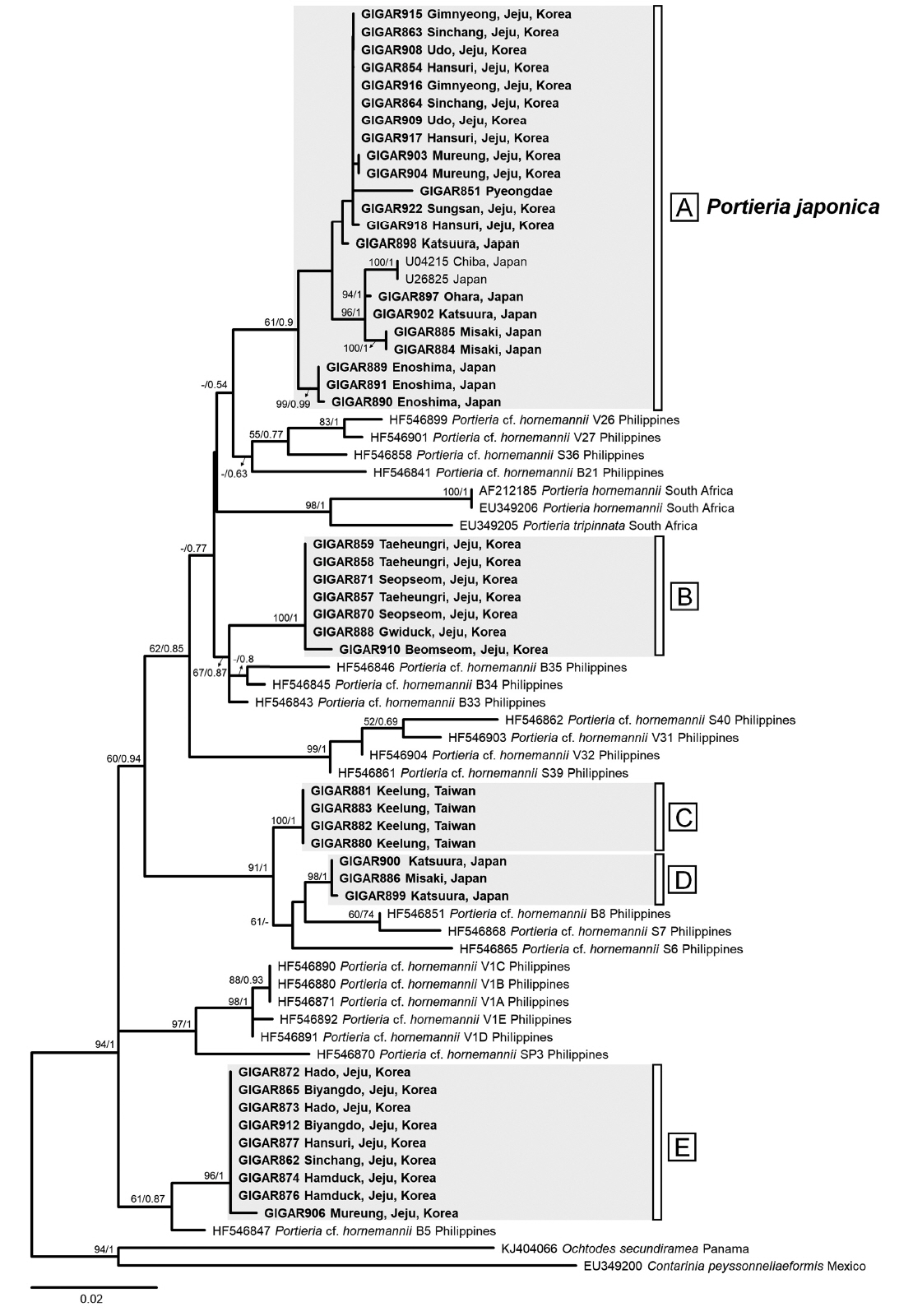
Our specimens from Korea, Japan, and Taiwan were divided into five lineages (A–E) (Fig. 1). All five lineages were reciprocally monophyletic, as supported by their high bootstrap values, except for lineage A (MLBt/BPP, A: 64/0.9, B: 100/1, C: 100/1, D: 98/1, E: 96/1); the sequence divergence was in the range of 2.0–5.8%. Interestingly, the partial sequences of Portieria cf. hornemannii from the Philippines did not match any of our specimens. Furthermore, the Philippine haplotypes were highly divergent, forming fragmented clades, each of which was associated with a different Portieria lineage revealed in this study.
Along with several specimens from Korea, most specimens from Japan, including those near the type locality of P. japonica and sequences of P. japonica in GenBank (U26825, U04215), were divided into lineage A, clarifying the designation of the P. japonica lineage in this study. The remaining specimens from this study were scattered into four separate lineages (B–E). The specimens from Taiwan and some from Japan formed two distinct lineages (C and D). Interestingly, those Korean specimens not grouped into the P. japonica lineage formed other two independent lineages (B and E). Since lineages B–E were identified as P. hornemannii based on their morphological similarity, we assumed them to be newly discovered cryptic species of P. hornemannii.
Fig. 2 showed representative images of morphology for the specimens from Korea, Japan, and Taiwan. Fig. 2A–C presents the P. japonica lineage, while the other images represent the cryptic lineages of P. hornemannii (Fig. 2D & E for lineage B, Fig. 2F for lineage C, Fig. 2G for lineage D, and Fig. 2H & I for lineage E). The most distinct difference among specimens of the P. japonica lineage and the others was the width of the thallus: the P. japonica lineage specimens usually had a relatively wide thallus (2–3 mm), but the other specimens of P. hornemannii assigned to the lineages B–E had much slender thallus (up to 1.2 mm in width). However, differentiating the P. hornemannii specimens into lineages B–E based solely on their external morphology was extremely difficult.
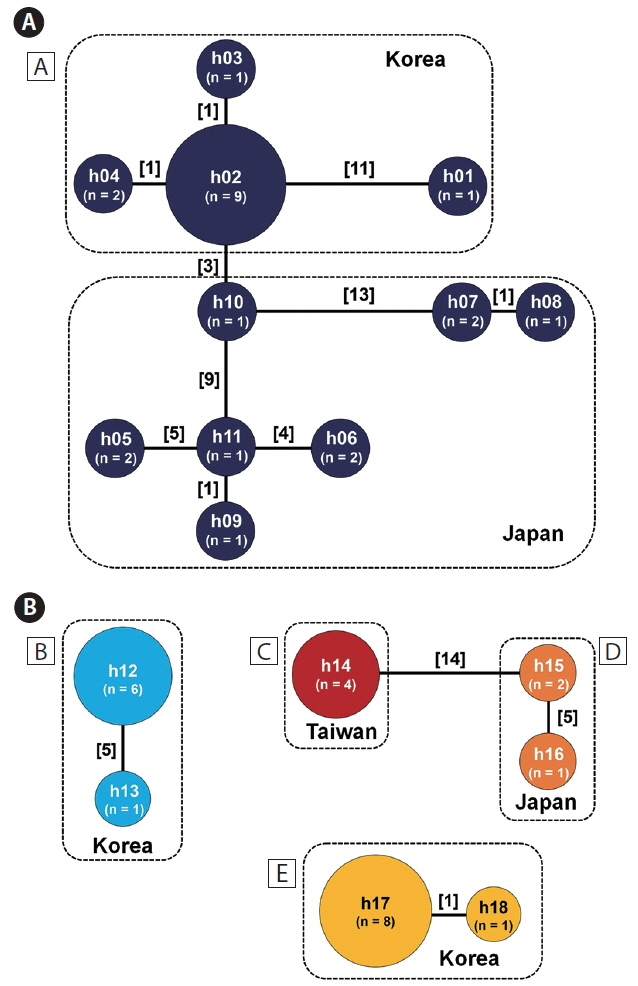
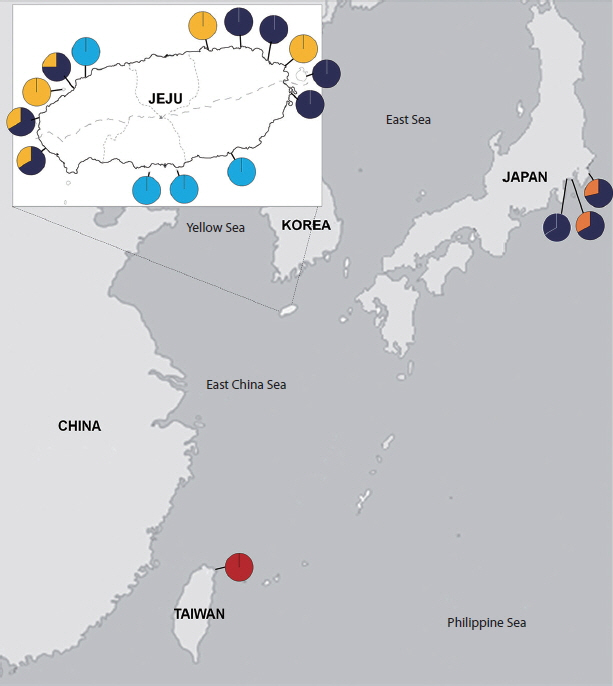
To understand the geographical structure within each lineage, we clustered 46 rbcL sequences (44 collected from this study + 2 sequences of P. japonica retrieved from GenBank) and constructed an association network (Fig. 3). We uncovered 18 unique haplotypes that could be grouped into lineages A–E and observed clear separations among the lineages. For instance, lineages B–E were each composed of specimens from single country and two or fewer haplotype clusters. However, lineage A was fragmented into 11 haplotypes, which were divided into two geographical subgroups from Korea and Japan. The result of cryptic lineage D suggested genetic segregation, even though it included only three samples (all from nearby regions in Japan). Interestingly, lineage C from Taiwan was distantly related to lineage D from Japan (Fig. 3). Overall, the geographical distribution of the lineages was indicative of sympatric distribution among the cryptic lineages of Portieria at several sites in Korea and Japan (Fig. 4). For instance, sympatry was observed between lineages A and E on the western coast of Jeju Island, Korea, as well as lineages A and D on the eastern coast of Japan.
DISCUSSION
Our rbcL data revealed five distinct lineages in the genus Portieria in Korea, Japan, and Taiwan. Lineage A, including specimens from Korea and Japan and the P. japonica sequences from GenBank, was identified as P. japonica, while the other lineages (B–E) could be considered as cryptic species of P. hornemannii. These results are in line with a molecular analysis of Portieria that found a number of cryptic species due to fine-scale endemism (Payo et al. 2013). This study also highlights the utility of molecular data to underpin cryptic species diversity particularly in morphologically indistinguishable groups. The finding of cryptic species of P. hornemannii in this study, with no matching to that from the Philippines, revealed the expansion of cryptic species diversity of P. hornemannii due to endemic environmental influences (Payo et al. 2013). However, this did not rule out the possibility of sympatry of phylogenetically distinct lineages that clearly revealed a sympatric distribution in the population analysis (Fig. 4).
Discriminating between P. japonica and P. hornemannii based on morphology alone is challenging. Although it relies mostly on the difference in width of the thallus and absence/presence of a sawtooth margin (Yoshida 1998), the cutoff value of thallus width that can be used to discriminate P. japonica from P. hornemannii is difficult to determine. In addition, the specimens identified as P. japonica in the study rarely showed sawtooth margins. This is further complicated by a lack of detailed descriptions of the morphological, vegetative, and reproductive development characteristics of P. japonica in contrast to those of P. hornemannii (Payo et al. 2011). After separating the specimens into lineage A–E based on the genetic evidence, the differences in thallus width between lineage A as P. japonica and other lineages corresponded well with the main diagnostic feature used to delimit P. japonica by Yoshida (1998).
Lineage A (P. japonica) was the most diverse in the rbcL haplotype network and divided into two geographical subgroups, Korean and Japanese (Fig. 3). The Japanese group was more genetically fragmented (7 haplotypes out of 10 samples) than the Korean group (4 haplotypes out of 13 samples). These two subgroups were closely connected each other by a 3-bp difference between h02 and h10, although there were high internal variations within the respective groups (a maximum 12-bp difference between h01 and h14 in the Korean group and maximum 28-bp difference between h08 and h05 in the Japanese group, respectively). This close connectivity between Korea and Japan (central Pacific Japan) was in contrast to the patterns observed in other gigartinalean species such as Chondracanthus intermedius Suringar (Hommersand) (see Yang and Kim 2016), Gloiopeltis complanata (Harvey) Yamada (see Yang and Kim 2018), and Chondrus ocellatus Holmes (see Hu et al. 2015), as well as other algal groups such as Gelidium elegans Kützing (see Kim et al. 2012) and Ishige okamurae Yendo (Lee et al. 2012), in which significant genetic breakages between the two regions had been detected.
The other lineages B–E were assumed to be cryptic species of P. hornemannii defined as genetically distinct but morphologically similar species. They are frequently identified as P. hornemannii because of their narrow thallus axes with many branches (Payo et al. 2011). However, the genetic diversity of P. hornemannii in the Philippines has demonstrated widespread local environmental influences and deep cryptic speciation within fine-scale areas (Payo et al. 2013), suggesting genetic differentiation of the species in other locations. Our results support the cryptic species diversity of P. hornemannii in the northwest Pacific region, despite insufficient sampling. The presence of hidden diversity in the genus Portieria is unsurprising given the multitude of examples exhibiting cryptic diversity, especially in a marine environment (Knowlton 1993). This study shows a high degree of genetic variation under morphological stasis in the specimens of P. hornemannii from the northwest Pacific.
Lineages B and E were only found in Jeju, Korea, which each contained one major and one minor haplotypes, respectively. Although a morphological similarity was confirmed between the two lineages, they were distantly related, showing 4.1–5.9% rbcL sequence divergence. The geographical distribution of two lineages demonstrates that species-level diversity could appear at even smaller scales on Jeju Island. Speciation at small geographical scales is often explained by the limited dispersal capacity of marine macroalgae with the absence of propagules (Payo et al. 2013). In addition, niche or physiological differences have been considered among coexisting cryptic macroalgal species (Zuccarello et al. 2001, Zuccarello and West 2003, Tronholm et al. 2010). Although lineages B and E were found together on Jeju Island, they were confined to separate site and lineage E cohabited with lineage A at several sites. Spatial partitioning at small scales appears to be a common mechanism allowing the coexistence of competitive species in marine environments (Wellenreuther et al. 2007). Recently, Tronholm et al. (2010) described the ecological differentiation between two species of brown algae, Dictyota from the Canary Islands, where pseudocryptic species have occupied distinct niches with respect to the exposure and timing of gamete release. These microhabitat differentiation has long been accepted as an important mechanism for the coexistence of algae (Noda 2009).
Lineages C and D were distributed in northern Taiwan and the central Pacific coast of Japan, respectively, separated by about 2,000 km. Lineage C in northern Taiwan was characterized by haplotype h14, which was connected to haplotypes h15 of lineage D from central Japan by 14 missing haplotypes. This association between lineages C and D, despite such a separation, can be partially explained by the Kuroshio Current, a warm current that arises from the North Pacific Equatorial Current flowing northeastwardly off the coast of Taiwan and Japan into the northern Pacific Ocean (Hu et al. 2011). However, the presence of 14 missing haplotypes between the northern Taiwan and the central Japan suggests long-term isolation in this regions. This is similar to the number of missing haplotypes in I. okamurae (brown alga) between Taiwan and other regions (Lee et al. 2012).
Intraspecific variation of ochtodene production in P. hornemannii was detected in Guam due to change in light exposure and temperature at different sites (Puglisi and Paul 1997). However, Matlock et al. (1999) demonstrated that the transplantation of P. hornemannii to a different site did not affect the mean concentration of apakaochtodenes A and B. They confirmed that simple environmental modifications did not alter the apakaochtodene levels in P. hornemannii. They also noted large differences in ochtodene concentrations among specimens within the same site. They determined that natural factors such as seasonal cycles of algal growth and the correlation between plant size and secondary metabolite production are affected the apakaochtodene contents of algae (Matlock et al. 1999). We cautiously speculate that the significant variability in secondary metabolites among sites and among specimens is due to different genetic populations of P. hornemannii, implying the existence of heterogeneous cryptic species. This is supported by the cryptic genetic diversity of P. hornemannii revealed in this study, whereby genetic isolation in different environmental regimes may have led to the physiological differentiation of these cryptic species. Additional studies will clarify the relationships between genetic and secondary metabolite variations in Portieria.