Abbreviations
BOLD
Barcode of Life Database
COI
cytochrome oxidase subunit I
MCE
Mesophotic Coral Ecosystems
NOAA
US National Oceanographic and Atmospheric Administration
NWHI
Northwestern Hawaiian Islands
PMNM
Papahānaumokuākea Marine National Monument
rbcL
ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit gene
INTRODUCTION
The peyssonnelioid genus Sonderophycus has a confusing taxonomic history. Denizot (1968) described the genus based on morphological differences between Peyssonnelia australis Sonder and other species in the genus Peyssonnelia. Sonderophycus was established with the combination Sonderophycus australis (Sonder) Denizot. Subsequently, Womersley and Sinkora (1981) established the genus Sonderopelta (based on S. coriacea Womersley & Sinkora) after examining the type specimen of P. australis Sonder in MEL and determining that it represented a species of Peyssonnelia. Thus, Sonderopelta coriacea Womersley & Sinkora was used to re-describe the taxon known as Sonderophycus australis (Sonder) Denizot. Further adding to the confusion, Silva et al. (1996) cited Peyssonnelia australis Sonder as a synonym of P. capensis Montagne. A phylogenetic analysis of the Peyssonneliales by Krayesky et al. (2009) demonstrated that the types of Sonderophycus (S. australis) and Sonderopelta (S. australis = P. capensis) formed a genus-level clade separate from Peyssonnelia based on rbcL and ribosomal large subunit sequence data, but in synonymizing the two genera they recognized the junior synonym, Sonderopelta. Wynne (2011, 2016) established that Sonderophycus had priority over Sonderopelta and established the type of the former as S. capensis (Montagne) M. J. Wynne.
The genus Sonderophycus is small, comprising only three currently recognized species: the type species, S. capensis (Montagne) M. J. Wynne, S. coriaceus (Womersley & Sinkora) M. J. Wynne, and S. fervens K. R. Dixon. Peyssonnelia caulifera Okamura is also a likely member and is not currently recognized as part of the genus only by virtue of a nomenclatural error in citation of the basionym when proposing the combination (Jeong et al. 2017, Guiry and Guiry 2020). The genus is distinctive from others in the Peyssonneliales in forming a well-defined clade in molecular phylogenetic analyses (Krayesky et al. 2009, Dixon and Saunders 2013, Dixon 2018), but also morphologically in the tomentum of fibrous rhizoids that anchor the thalli to the substratum, the lack of hair cells, and the tendency of lower perithallial filaments to be sharply angled (Denizot 1968, Jeong et al. 2017, Dixon 2018).
Studies of the peyssonnelioid algae of the Hawaiian Islands are relatively few. Four species of Peyssonnelia have been reported from Hawai‘i, namely, P. conchicola Piccone & Grunow, P. inamoena Pilger, P. rubra (Greville) J. Agardh by Abbott (1999), and P. japonica (Segawa) Yoneshigue by Kato et al. (2009). A number of sequences of Peyssonneliales were presented in Sherwood et al. (2010), but the species-level taxonomy of peyssonnelioid specimens in Hawai‘i was not investigated in further detail at that time. The genus Ramicrusta was recently reported for the first time from the Hawaiian Islands, with the proposal of two (presumably) endemic species; R. hawaiiensis A. R. Sherwood and R. lehuensis A. R. Sherwood (Sherwood et al. in press). Thus, a total of six peyssonnelioid species have been reported from Hawai‘i, representing Peyssonnelia and Ramicrusta; no species of Sonderophycus or its synonyms have been previously recorded.
This study is part of the ongoing effort to characterize the mesophotic algae of the Hawaiian Islands. During mesophotic expeditions to Papahānaumokuākea Marine National Monument (PMNM) led by the US National Oceanic and Atmospheric Administration (NOAA) in 2015 and 2016, three specimens attributable to the genus Sonderophycus were collected. These algae were observed as a dominant member of the benthic community at Kure Atoll, suggesting an ecologically important role for this species. Molecular and morphological characterization of these specimens is here used to support the proposal of these specimens as a new species of Sonderophycus.
MATERIALS AND METHODS
Three specimens of Sonderophycus were collected in 2015–2016 by technical rebreather diving as part of marine survey expeditions to Kure Atoll, PMNM in the Northwestern Hawaiian Islands (NWHI), between 88 and 92 m depth (Table 1). Field work was conducted under PMNM permit PMNM-2015-029 to R. Kosaki. Specimens were pressed as herbarium vouchers for morphological analysis and preserved in silica gel desiccant for molecular characterization. Specimens were examined and cleaned under a Zeiss SteREO Discovery v12 dissecting microscope (Pleasanton, CA, USA). Morphological and anatomical investigations of the samples were conducted by rehydrating small pieces of the specimens in a detergent (Modified Pohl’s Solution, Clark unpublished data: https://www.eeob.iastate.edu/research/bamboo/pdf/anatomy_protocols.pdf) for 30 min, decalcifying small pieces of attached coral rubble in 5% HCl for 15 min, hand sectioning with a double-edged razor blade, staining with 1% aniline blue, and mounting in 30–50% Karo. Photomicrographs were taken on a Zeiss AxioImager A1 compound light microscope (Pleasanton, CA, USA) with an Infinity2-1RC digital camera (Lumenera Corporation, Ottawa, ON, Canada).
Specimens were extracted for genomic DNA using an OMEGA E.Z.N.A. Plant DNA DS Kit (OMEGA Biotek, Norcross, GA, USA). The cytochrome oxidase subunit 1 (COI) DNA barcode marker (658 bp) was amplified using the GazF1 and GazR1 primers (Saunders 2005). The rbcL (ribulose-1,5-bisphosphate carboxylase / oxygenase large subunit, 1,335 bp) marker was amplified as three overlapping fragments using the following primer pairs: F7 and R753 (Gavio and Fredericq 2002), F492 (or alternatively F577) and R1150 (Freshwater and Rueness 1994), and F993 and R-rbcS start (Freshwater and Rueness 1994). Successful PCR products were submitted for sequencing to the University of Hawai‘i at Mānoa Advanced Studies for Genomics, Proteomics, and Bioinformatics core facility or through GENEWIZ (South Plainfield, NJ, USA). Raw sequence reads for each gene were assembled, edited, and aligned using the MUSCLE v. 3.8.425 plug-in (Edgar 2004) in Geneious Prime 2019.1.3 (http://www.geneious.com) with other available sequences for Sonderophycus and related genera from GenBank and the Barcode of Life Database (BOLD) (Supplementary Table S1). DNA barcode analysis of the COI sequences was performed by constructing a neighbor-joining framework based on Kimura-2-parameter distances using MEGA X (Kumar et al. 2018). For the rbcL phylogenetic analyses, newly generated sequences were aligned with reference sequences and analyzed with PartitionFinder v. 1.1.1 (Lanfear et al. 2012). Maximum likelihood analyses were performed on all alignments using RAxML-HPC2 on XSEDE v. 8.2.10 (Stamatakis 2014) via the CIPRES gateway (Miller et al. 2010) with 1,000 bootstrap replicates, and using the GTRCAT model. Bayesian inference was performed using the MrBayes plug-in v. 3.2.6 (Huelsenbeck and Ronquist 2001) through Geneious Prime 2019.1.3 (http://www.geneious.com) using four chains of Metropolis-coupled Markov Chain Monte Carlo for 1,000,000 generations and sampling every 100 generations; 100,000 chains were removed as burn-in prior to determining posterior probabilities.
RESULTS
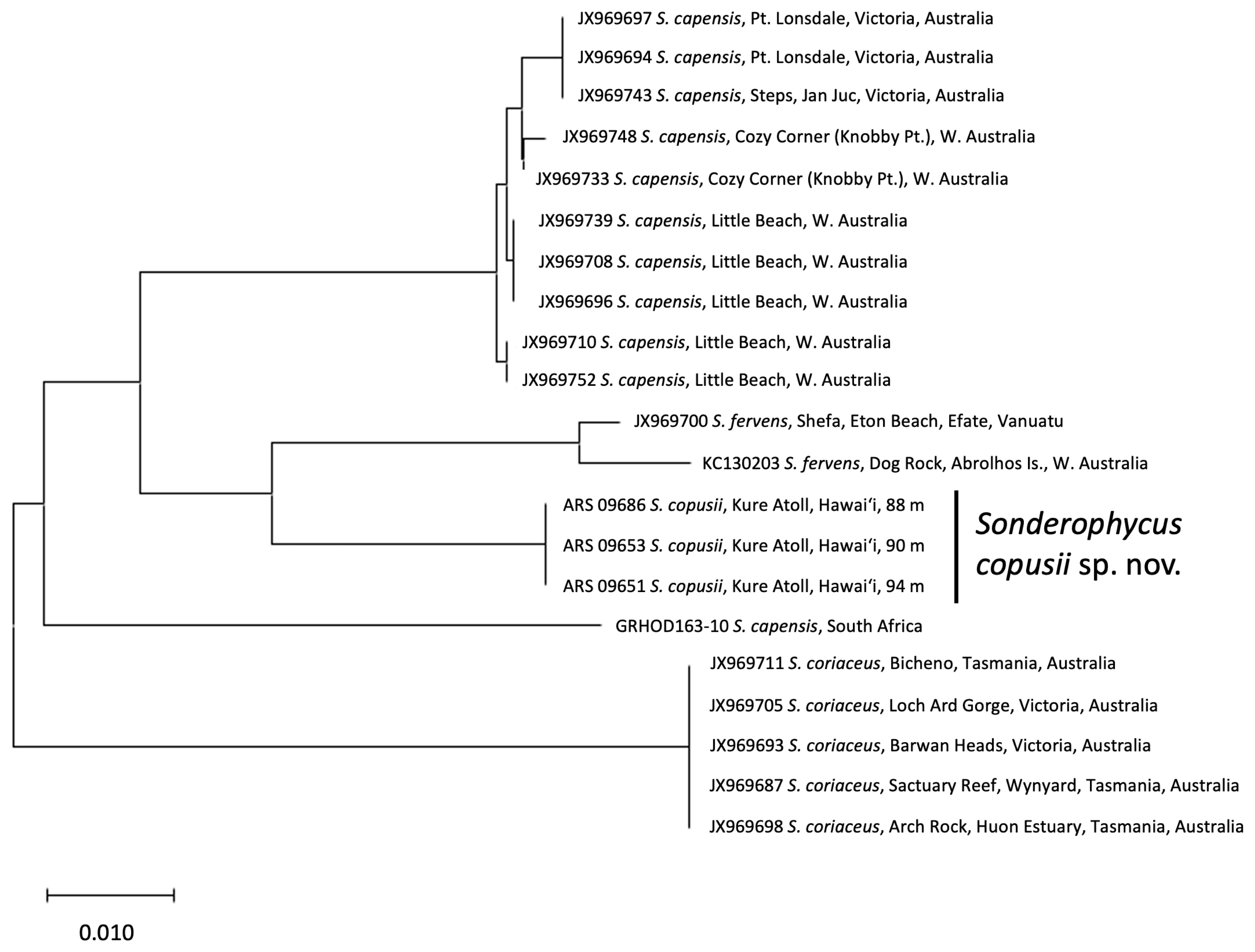
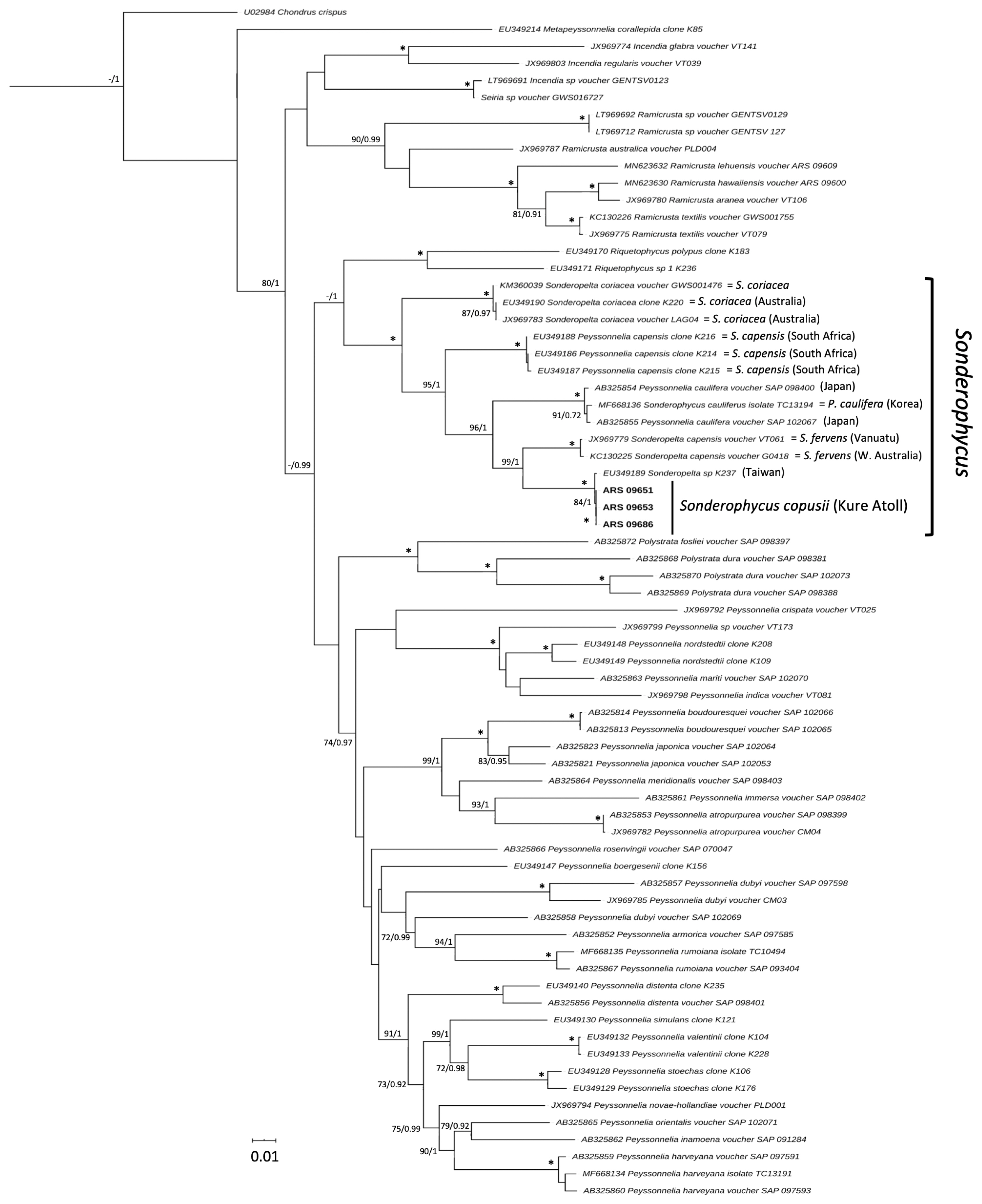
DNA barcoding analysis of COI sequences of the three new Hawaiian Sonderophycus specimens and 18 additional specimens of Sonderophycus representing S. capensis, S. coriaceus, and S. fervens clearly indicated that the Hawaiian specimens were identical to one another yet distinct from other species of the genus (Fig. 1). Of the currently recognized and likely members of the genus, only P. caulifera was not represented in the COI barcoding analysis. All taxa represented in the analysis include specimens from, or close to, their type locality. The Hawaiian specimens shared 95.0–95.1% identity with their most similar congeners (S. fervens; JX969700, KC130203). In addition, COI sequence data for S. capensis clearly distinguished specimens from South Africa (type locality = Durban, South Africa) and Australia, demonstrating that the Australian material is likely a distinct species from S. capensis. Phylogenetic analysis of rbcL sequences representing all known members of Sonderophycus plus P. caulifera, as well as other representatives of the Peyssonneliales, yielded a well-resolved phylogeny that indicated full support for the monophyly of the genus Sonderophycus (Fig. 2). Clades corresponding to S. capensis, S. coriaceus, S. fervens, P. caulifera, and the Hawaiian specimens plus one additional GenBank sequence (see below) were all resolved with full support. The Hawaiian specimens characterized in this study were allied with a sequence from Magang Harbor, Taiwan, at 4 m depth (EU349189), differing from this sequence by only two nucleotides (Fig. 2). The closest relative of the Hawaiian Sonderophycus is the Australian taxon, S. fervens.
The Hawaiian specimens are here recognized as a new species of Sonderophycus based on the molecular distinctiveness of the specimens within the context of other species of the genus, as well as on morphological characters, as outlined below.
Sonderophycus copusii A. R. Sherwood sp. nov. (Fig. 3A–J)
Description.
Thallus encrusting with the margins free, typically on rock or coral rubble, attached by rhizoids. Deep brick red to dark dusky rose in color, radial striations, and faint concentric rings visible on dorsal surface. Thallus composed of a single, broadly lobed blade, orbicular in shape, and cartilaginous, reaching 8.5 cm in diameter. Cystoliths and hypobasal calcification present. Blades 65–210 μm thick, composed of 6–10 cell layers. Hypothallus composed of radiating and parallel filaments, cells 8–9 μm diameter × 11–18 μm length. Occasional secondary perithallial growth. Perithallial filaments arising at approximately 45° to the hypothallus, but straightening to upright (90°) near the dorsal surface of the thallus. Perithallial cells elongate in shape toward basal region of perithallus, 10–12 × 30–45 μm (diameter × length), and closer to isodiametric in upper region of perithallus, 8–13 × 6–16 μm (diameter × length). Rhizoids multicellular, 8–15 μm μm diameter, long and fibrous, tangled, felt-like, and cut off at an acute angle from the distal end of the hypothallus towards the growing margin; on emergence from the hypobasal cuticle the angle sometimes lessens; cuticle 10–13 μm thick. Hair cells absent. Tetrasporangia 15–20 μm diameter and 50–80 μm long, cruciately divided, borne on paraphyses produced in elevated, subcircular gelatinous nemathecia (500–1,200 μm diameter) on thallus surface. Gametangial reproduction not observed.
Holotype
BISH 776116 (ARS 09653, Kure Atoll, Hawai‘i, 28.49845, −178.341712, depth 90 m, Jun 5, 2016, leg. J. Hansen & B. Hauk [NWHI-543], sheet 1). Tetrasporangial plants.
Etymology
Named for Joshua Copus, our colleague and friend, and a doctoral student at the University of Hawai‘i at Mānoa who passed away on Nov 12, 2019, while diving during a research expedition to study the mesophotic coral reefs of the Solomon Islands.
Distribution
Kure Atoll, Papahānaumokuākea Marine National Monument, Hawai‘i, USA, from 88–94 m depth.
DNA sequence data
GenBank accessions MT012464, MT012466, MT012468 (COI) and MT012465, MT012467, MT012469 (rbcL).
Habit and morphology
Dense beds of Sonderophycus copusii were photographed at Kure Atoll, with the species covering more than 50% of the substratum. In cross-section, this taxon strongly resembles other members of the genus, with perithallial filaments arising at an angle of approximately 45° from the hypothallus. Specimens also have an obvious, protruding marginal apical cell at the thallus margin, which corresponds to reports of other species of Sonderophycus. The rhizoids of S. copusii form a remarkably thick tomentum, which are frequently entangled with fungal hyphae. Cystoliths are evident in material that has not been decalcified. Although male and female gametophytes were not observed, tetrasporangial plants were among those characterized.
Key to the species of Sonderophycus (including P. caulifera)
DISCUSSION
Dixon (2018) remarked that Peyssonnelia and Sonderophycus cannot be unequivocally distinguished based on morphology despite some characters appearing consistent within Sonderophycus, and that molecular phylogenetic analyses provide the strongest evidence for the monophyly of Sonderophycus within a polyphyletic Peyssonnelia. Recognition of peyssonnelioid genera in recent years through combined molecular and morphological analyses has led to the description of several new genera (e.g., Incendia and Seiria) (Dixon and Saunders 2013, Dixon 2018), the clarification of others that have been recognized for some time (e.g., Polystrata, Ramicrusta, Sonderophycus) (Zhang and Zhou 1981, Kato et al. 2006, Krayesky et al. 2009, Pueschel and Saunders 2009, Ballantine et al. 2016), and the reinstatement of others (e.g., Ethelia, which was recently reinstated as distinct from Polystrata and placed into its own family) (Dixon et al. 2015). This theme of phylogenetic clarification followed by intensive taxonomic overhaul has been common across the red algae in recent decades (e.g., recognition of many new and resurrected genera within the Batrachospermales, Gracilariales, Kallymeniaceae) (e.g., Entwisle et al. 2009, Saunders et al. 2017, Gurgel et al. 2018), and is an indication of the taxonomic revival that is currently underway in the field of phycology (De Clerck et al. 2013).
Recognition of the Hawaiian specimens as a new species, Sonderophycus copusii, is supported by both morphological and molecular phylogenetic comparisons, and sequence data for the rbcL and COI markers clearly distinguish the species from all other described taxa in the Peyssonneliales. However, no single morphological character is diagnostic for S. copusii, and the distinction of this species from others in the genus relies on a combination of features: the presence of occasional secondary perithallial growth, emergence of rhizoids from the hypobasal cuticle at a strongly bent angle, a lack of horizontally directed filaments in the lower perithallus, and the lack of a stipe. Secondary perithallial growth was reported for S. fervens (Dixon 2018) and was occasionally observed in S. copusii specimens but is not known in other species of Sonderophycus. The emergence of the rhizoids from the hypobasal cuticle at a strongly bent angle appears to be a character shared by several species of Sonderophycus (S. capensis, S. copusii, and S. fervens), while the presence of a stipe is known only in S. coriaceus (which is prominently three-dimensional, with thallus lobes raised from the substratum) (Womersley and Sinkora 1981, Dixon 2018). Sonderophycus copusii is similar to both S. fervens and Peyssonnelia caulifera; S. fervens and S. copusii are distinguished by the former having prominent radial streaks on the thallus surface, which are present but not prominent in the latter, and the lack of horizontally directed filaments in the lower perithallus (as per Dixon 2018) in S. copusii. Similarly, S. copusii differs from P. caulifera in having acutely bent rhizoids at the point of emergence from the hypobasal cuticle (Dixon 2018).
The Sonderophycus copusii specimens reported in the current study are almost identical in rbcL sequence (differing by only 2 bp) to a specimen published by Krayesky et al. (2009) (as “Sonderopelta sp. K237”), which was collected at 4 m depth in Magang Harbor, Taiwan. Interestingly, the Hawaiian S. copusii specimens were collected from mesophotic depths (the taxon is known only from 88–94 m depth), and thus far are limited in distribution to the Hawaiian Islands. Given the lack of comparative morphological data for the Taiwanese specimen, it is unknown whether the Hawaiian and Taiwanese specimens represent the same species, although they are certainly very closely related, and their close relationship may shed light on the biogeographic affinities of the Hawaiian mesophotic Sonderophycus. Molecular phylogenetic analyses that have been conducted to date on the biogeographic patterns of Hawaiian mesophotic algae have started to yield insight into the biogeographic affinities of the flora. For example, one of four new sea lettuces was closely allied with an undescribed taxon from the Kermadec Islands and western Australia, two were closely associated with specimens from Mage Island, Japan, and the fourth species was not closely associated with any other known sea lettuce species (Spalding et al. 2016). A recent analysis of the red algal genus Martensia from Hawai‘i revealed the presence of five species, all with mesophotic representation, four of which were newly described in the study and presumed endemic to the Hawaiian Islands, and one that was demonstrated to be a new record for the islands (M. albida Y. Lee) and previously known only from Korea (Sherwood et al. 2019). Two new species described in the peyssonnelioid genus Ramicrusta from Lehua Island, Hawai‘i (one from mesophotic depths) were distinct from all other described species thus far but were allied with taxa known from the western Pacific / Australasia and the Caribbean (Sherwood et al. in press). Thus, the close association of S. copusii with a Taiwanese specimen is consistent with the majority of known biogeographic associations of Hawaiian mesophotic algae to taxa from the Asia-Pacific region. As the species-level taxonomy of the Hawaiian mesophotic flora is clarified in the coming years, our understanding of the biogeographic associations of the flora will also be resolved in greater detail.
The beds of Sonderophycus copusii from Kure Atoll are consistent with the Corallinales / Peyssonneliales assemblages noted in Mesophotic Coral Ecosystems (MCEs) of the Caribbean, Mediterranean, and the Red Sea, and may be part of a broader pattern of dominant assemblages at depth in these ecosystems. The Kure Atoll Sonderophycus beds bear similarity to the “Peyssonnelia beds” reported from deep waters (30–163 m depth) of various areas of the Mediterranean, which are reported to be patchy in distribution (Augier and Bourdouresque 1978, Ballesteros 1994). Similarly, Aponte and Ballantine (2001) reported mesophotic peyssonnelioids abundant from 60–120 m in the Bahamas, and as part of dominant assemblages in that island group. Likewise, Dullo et al. (1990) noted dominant Corallinales / Peyssonneliales assemblages to 110 m in deep forereef environments in the Red Sea. The ecological role and significance of these mesophotic peyssonnelioid beds is unknown, but it has been hypothesized that they may contribute substantially to sediment production (Ballesteros 1994).
The genus Sonderophycus has a relatively broad, pantropical-warm temperate distribution, but with the majority of species known from the Pacific (Guiry and Guiry 2020). Sonderophycus capensis has by far the broadest reported distribution of all the species in the genus, with its distribution including Africa, the Indian Ocean, Asia, Australia, and New Zealand, the Pacific Islands, South America, and the Red Sea (e.g., Guimarães and Fujii 1999, Kang et al. 2006, Guiry and Guiry 2020). Peyssonnelia caulifera is moderately broad in distribution, with reports from Asia and southeast Asia, and the Pacific Islands (Guiry and Guiry 2020). The remaining species have much more restricted distributions, with S. fervens known from northwest Australia, Indonesia, the Philippines, and Vanuatu, S. coriaceus known from south Australia and Tasmania, and S. copusii known only from the Northwestern Hawaiian Islands. The center of diversity for the genus is clearly in the Pacific, with all species known from this ocean basin, and only one species (S. capensis) known outside it (Guiry and Guiry 2020).
Studies of the biodiversity of MCEs have indicated some preliminary trends that will be interesting to explore as more of the algal diversity in these deep-water environments is characterized. For example, fishes have been shown to generally have narrower geographic distributions than their shallow counterparts, high levels of endemism are suggested for all groups of organisms in Hawaiian MCEs, and the degree of shallow-to-deep connectivity has been shown to vary by taxon (Pyle and Copus 2019). Combined with the impressive diversity of peyssonnelioid species that have been described in recent years (e.g., Pueschel and Saunders 2009, Ballantine and Ruíz 2010, 2011, 2012, Dixon and Saunders 2013), it is clear that future contributions to mesophotic algal systematics and taxonomy will be important for understanding not only the floristic composition of individual regions, but to clarify the interpretation of these broader patterns of diversity in the mesophotic.