Abbreviations
AAPH
2,2′-azobis(2-amidinopropane) hydrochloride
ABTS
2,2′-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid)
ACN
acetonitrile
CPC
preparative centrifugal partition chromatography
DCFH2
2,7-dichlorodihydrofluorescein
DCFH-DA
2′,7′-dichlorodihydroflurescin diacetate
DMEM
Dulbecco’s modified Eagle medium
dpf
days post-fertilization
DW
distilled water
FBS
fetal bovine serum
GAE
gallic acid equivalent
HPLC
high-performance liquid chromatography
LDH
lactate dehydrogenase
MTT
3-(4-5-dimethyl-2yl)-2-5-diphynyltetrasolium bromide
NMR
nuclear magnetic resonance
PBS
phosphate buffered saline
ROS
reactive oxygen species
SPE
70% ethanol extracts of Spirogyra sp.
SPEC
chloroform fraction of SPE
SPEE
ethyl acetate fraction of SPE
SPEH
hexane fraction of SPE
SPEW
water fraction of SPE
UV
ultraviolet
INTRODUCTION
Although ultraviolet (UV) is essential for human life, it can impair the ability of basal keratinocytes to maintain skin homeostasis against UV-induced damage, by which can be a major contributor in the development of skin cancers (Kong et al. 2015). Ultraviolet B (UVB), a type of UV in the wavelength ranging from 280 nm to 315 nm, has been studied in the epidermal sun-burn of keratinocytes that are destined to DNA damage and apoptosis after sun exposure. Although humans are exposed to UVA 10 to 100 folds more than UVB, UVB is known to be 1,280 folds more effective at inducing erythema, thus more than 90% of erythema induced by UV irradiation is attributed to UVB (Ryu et al. 2015). Furthermore, due to the ozone layer becoming thinner from anthropogenic activities, an excessive amount of UVB in the biosphere is being irradiated (Liu et al. 2015). UVB stimulates generation of reactive oxygen species (ROS), including superoxide anion radical (O2−), hydrogen peroxide (H2O2), hydroxyl radical (HO·), and singlet oxygen (1O2) that may result in the destruction of keratinocytes through cellular damage and apoptosis (Kulms and Schwarz 2002, Rezvani et al. 2006, Kim et al. 2007, Gill and Tuteja 2010, Oh et al. 2016, Sanjeewa et al. 2016).
Many studies have been conducted to find the bioactive extracts or active ingredients derived from natural sources, particularly having polyphenol structure including hydroxyl groups on aromatic rings, which can modulate the generation of UVB induced-ROS (Samarakoon et al. 2013, Kim et al. 2014, Farooqi et al. 2015, Lee et al. 2015b, 2015c, 2015d, Alves et al. 2016, Fernando et al. 2017a, 2017b).
Spirogyra sp. is the freshwater green alga used as a biosorbent to remove heavy metal ions from wastewaters (Gupta et al. 2001, Bishnoi et al. 2005, Gupta et al. 2006, Mohan et al. 2007, Gupta and Rastogi 2008). Spirogyra sp. has recently been known for its pharmacological activities as an anti-hypertension, resulted from its isolated gallic acid (Kang et al. 2015), as well as its potential as a bioenergy producing resource (Eshaq et al. 2010, Ramaraj et al. 2015). Our previous study investigated the protective effect of Spirogyra sp. against 2,2′-azobis(2-amidinopropane) hydrochloride (AAPH)-induced lipid peroxidation in zebrafish (Lee et al. 2015a). However, the protective effects of Spirogyra sp. against UVB-induced apoptosis have not yet been assessed.
Zebrafish (Danio rerio) has been increasingly used as an in vivo animal model for assessment of drug efficacy, toxicity and safety (Hwang et al. 2016). This is due to its short life span, comparatively small size, and similarity to mammals (Ko et al. 2011). Zebrafish have previously been used in several studies as an in vivo model for UV protective research (Dong et al. 2007, Ko et al. 2011, Yang et al. 2012). Hence, we selected zebrafish as an in vivo model to investigate the UVB protective effect of Spirogyra sp. in this study.
In this study, we have evaluated the phenolic content of 70% ethanol extract of Spirogyra sp. by three stage serial fractionations and examined the effects of gallic acid derivatives of Spirogyra sp. against UVB-induced ROS production and apoptosis in in vitro HaCaT cells and in vivo zebrafish. Furthermore, we isolated the bioactive compounds from the gallic acid derivatives by preparative centrifugal partition chromatography (CPC). The purpose of the present work was to assess the potential of the gallic acid derivatives of Spirogyra sp. in prevention of UVB-induced skin damage and to isolate its bioactive compounds.
MATERIALS AND METHODS
Chemicals and reagents
The fluorescent probe 2′,7′-dichlorodihydroflurescin diacetate (DCFH-DA) and 3-(4-5-dimethyl-2yl)-2-5-diphynyltetrasolium bromide (MTT), dimethyl sulfoxide and phosphate buffered saline (PBS), standard gallic acid, and 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid) (ABTS) were purchased from Sigma Co. (St. Louis, MO, USA). Online high-performance liquid chromatography (HPLC)-grade acetonitrile (ACN) was purchased from Burdick & Jackson (Muskegon, MI, USA). The Dulbecco’s modified Eagle medium (DMEM), penicillin/streptomycin, and fetal bovine serum (FBS) were purchased from Sigma Co. The solvents used for CPC were purchased from Fluka Co. (Buchs, Switzerland). All other chemicals used in this study were of analytical grade.
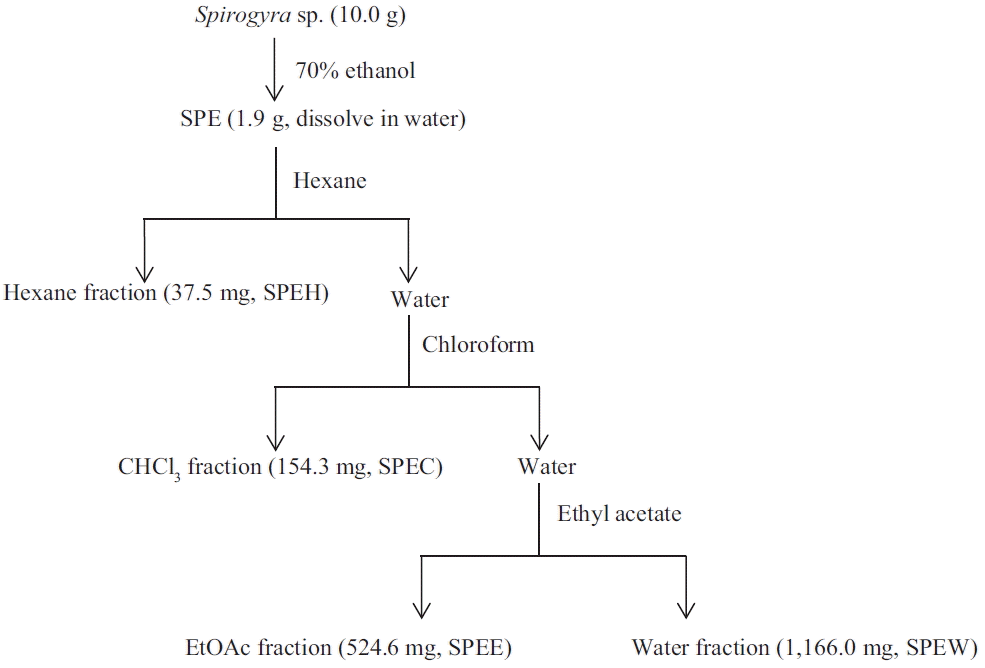
Fractionation and characterization of 70% ethanol extracts of Spirogyra sp. (SPE)
Fractionation
Spirogyra sp. was collected in the spring season, in January 2014, from the shallow ponds of Kongju, South Korea (36°20′34″ N, 127°12′28″ E). Lyophilized algae powder (10 g) was extracted by 70% ethanol (1 L) under continuous shaking at 120 rpm for 24 h at 25°C without light and subsequently filtered through Whatman No. 4 filter paper. The extracts were concentrated using rotary evaporator at a low pressure and temperature less than 37°C. Finally, we obtained the 70% ethanol crude extract of Spirogyra sp. that is referred to as SPE. SPE was further fractionated by different polarity organic solvents that include hexane, chloroform, and ethyl acetate. The hexane, chloroform, ethyl acetate, and water fractions of SPE were named as SPEH, SPEC, SPEE, and SPEW, respectively.
Phenolic content measurement
The total phenolic contents of SPE and its fractions were determined based on the method described by Chandler and Dodds (1983). Gallic acid was used as the reference phenolic compound to construct the standard curve and calculate the phenolic content of samples.
Determination of ABTS radical scavenging activity of SPEE by HPLC system
The ABTS radical scavenging activity of SPEE was analyzed by online HPLC system (Lee et al. 2015a). A concentration of 2 mM ABTS radical solvent containing 2.5 mM potassium persulfate was prepared and incubated with samples for 24 h for radical stabilization. HPLC system equipped with binary Waters 515 pump, Waters 2489 UV/Vis and 2998 photodiode array detector, and Waters 2707 auto-sampler, coupled with the interface ABTS+ radical analyzer (Waters, Mailford, MA, USA) was used. A volume of 10 μL SPEE (5 mg mL−1, dissolved in methanol) were injected to an Atlantis T3 3 μm 3.0 × 150 mm column (Waters) and run at 0.3 mL min−1 flow rate using a gradient of ACN and distilled water (DW) solvent system: 0–10 min, 0% ACN and 100% DW; 10–30 min, 25% ACN and 75% DW; 30–80 min, 50% ACN and 50% DW; 80–90 min, 100% ACN and 0% DW; 90–110 min, 100% ACN and 0% DW. Wavelength was at 220 nm. SPEE was eluted through the column, and the elution was reacted with ABTS radical in the reaction coil at 40°C. The absorbance of the reaction mixture was measured as the negative peak by UV/Vis detector at a wavelength of 680 nm.
Separation and identification bioactive compounds of SPEE
The active compounds from SPEE were separated in CPC chromatography (Sanki Engineering, Tokyo, Japan), and the target fraction was further purified by HPLC following above the methods until its amount and purity were enough for structural identification. The structure of target compound was identified by the spectroscopic data provided by the nuclear magnetic resonance (NMR) spectroscopy. The 1H-NMR and 13C-NMR spectrums were recorded on JEOL DELTA-400 (400 Hz) spectrometer (Jeol, Tokyo, Japan). The chemical shifts of the spectrums were reported as δ relative to tetramethylsilane as an internal standard.
In vitro analyses
Cell culture and UVB irradiation
Human keratinocytes (HaCaT cells) were purchased from Korean Cell Line Bank. The HaCaT cells were maintained in DMEM supplemented with 10% heat-inactivated FBS, streptomycin (100 μg mL−1), and penicillin (100 unit mL−1) at 37°C with 5% CO2. Cells were sub-cultured at 3 day intervals and seeded at a density of 1.0 × 105 cells per well in a 24-well plate. UVB irradiation was imposed using a UVB meter (UV Lamp, VL-6LM; Vilber Lourmat, Torcy, France) with a fluorescent bulb emitting 280–320 nm wavelengths with a peak at 313 nm. HaCaT cells were irradiated at a dose of 30 mJ cm−2 of UVB in PBS (Heo et al. 2010). Cells were subsequently incubated until analysis.
Determination of cell viability and apoptotic cell death analysis
For analyzing the protective effect of SPEE against UVB-induced cell damage, HaCaT cells were treated with different concentrations of SPEE (25, 50, and 100 μg mL−1) for 2 h at 37°C. Cells were then exposed to 30 mJ cm−2 of UVB and incubated for another 24 h. Subsequently, cell damage and proliferation were assessed by lactate dehydrogenase (LDH) release assay and MTT assay respectively as described previously (Kang et al. 2014).
The protective effect of SPEE against apoptosis was evaluated by nuclear staining and cell cycle analysis as per manufacturer’s protocol (Wijesinghe et al. 2013). The stained cells were observed using a fluorescence microscope (Olympus, Tokyo, Japan) to examine apoptotic body formation. Histograms of cell distribution in the different cell cycle phases were analyzed by the Quest and Mod-Fit (BD Bioscience, San Jose, CA, USA). The anti-apoptotic activity of SPEE was evaluated by assessing the change in the percentage of cell distribution at sub-G1 phase.
Determination of intracellular ROS generation induced by UVB irradiation in HaCaT cells
Subsequent to 30 min incubation with SPEE, cells were labeled with DCFH-DA (500 μg mL−1) for 30 min and exposed to UVB irradiation (30 mJ cm−2). Fluorescent intensity was determined at an excitation wavelength of 485 nm and an emission wavelength of 535 nm, using a fluorescent microplate reader (BioTek, Synergy HT, Wonooski, VT, USA).
In vivo analyses
Origin and maintenance of parental zebrafish
The adult zebrafish were purchased from a commercial dealer (Seoul aquarium, Korea). Fish were separately kept in 3 L acrylic tanks at 28.5°C, with a 14 : 10 h light : dark cycle. Zebrafish were fed 3 times day−1, 6 days wk−1, with Tetramin flake food supplemented with live brine shrimps. Embryos were obtained from natural spawning that was induced in the morning by turning on the light. Collections of embryos were completed within 30 min.
Determination of UVB protective effect of SPEE against UVB-irradiation in vivo zebrafish
The 2-days post-fertilization (dpf) embryos were used to evaluate the UVB protective effect of SPEE in this study. Embryos were collected in 12-well plates (10 embryos per well) containing 2 mL embryo medium, and subjected to SPEE with a final concentration of 25, 50, and 100 μg mL−1 for 1 h. The embryos were then washed with fresh media and exposed to 50 mJ cm−2 UVB per individual (Heo et al. 2009). The exposed embryos to UVB were transferred into a 24-well plate and labeled with DCFH-DA (400 μg mL−1) for 1 h. The embryos were washed twice with fresh media and anesthetized using phenoxyethanol prior to visualization (Ko et al. 2011). The anesthetized embryos were photographed under a microscope equipped with Cool SNAP-Pro color digital camera (Olympus). Fluorescence intensities were quantified by an image J program.
Statistical analysis
All the experiments were performed in triplicate. The data were expressed as the mean ± standard error (SE), and one-way ANOVA test (using Statistical Product and Service Solutions 11.5 statistical software) was used to statistically compare the mean values of each treatment. Significant differences between the means of parameters were determined by Duncan’s multiple range tests, p < 0.05 and p < 0.01 were considered as significantly different.
RESULTS
Phenolic content for serial fractions of SPE
The 70% ethanol extract of Spirogyra sp. was prepared and 1.92 g of it was used for serial fractionations using solvents of increasing polarity (Fig. 1). Serial fractionation yielded various quantities, with hexane fraction being 37.50 mg, followed by a chloroform fraction, an ethyl acetate fraction, and a water fraction yielding 154.30, 524.61, and 1,166.00 mg of residue, respectively (Table 1).
Each fraction was assessed for its total phenolic content. As shown in Table 1, total phenolic contents of the different fractions from serial fractionations ranged from 70.74 ± 2.61 to 774.42 ± 2.61 mg g−1 gallic acid equivalent (GAE). Through serial fractionations, phenolic content of SPE (260.47 ± 5.21 mg mL−1 GAE) enhanced 2.97 times to 774.42 ± 2.61 mg g−1 GAE of SPEE (Table 1).
Online HPLC-ABTS analysis of SPEE
Online HPLC-ABTS analysis, utilizing parallel chemical-detection methods, simultaneously separated and quantified the active peaks. The online analysis of SPEE exhibited 8 main peaks at the wavelength of 290 nm, corresponding to 8 ABTS radical scavenging peaks (Fig. 2A).
The online HPLC-ABTS data were assessed from the point that the area under the negative peak producing antiradical chromatogram (obtained at 680 nm) could show the total antiradical capacity of the sample analyzed. The areas of chromatograms at 680 nm were integrated for the 0–35 min time range to express total activity. Comparing the antiradical capacity of different peaks, 70% or above of total antiradical activity of the individual peaks were determined as 1, 5, and 8 peaks with higher on-line antioxidant activities. Peaks showing higher antiradical activities were identified as gallic acid, ethyl gallate, and methyl gallate at CPC chromatogram (Fig. 2B). In particular, ethyl gallate (C9H10O2, 198.17 g mol−1) was a newly identified and isolated compound from Spirogyra sp., given the details of 1H and 13C-NMR (see Supplementary Table S1).
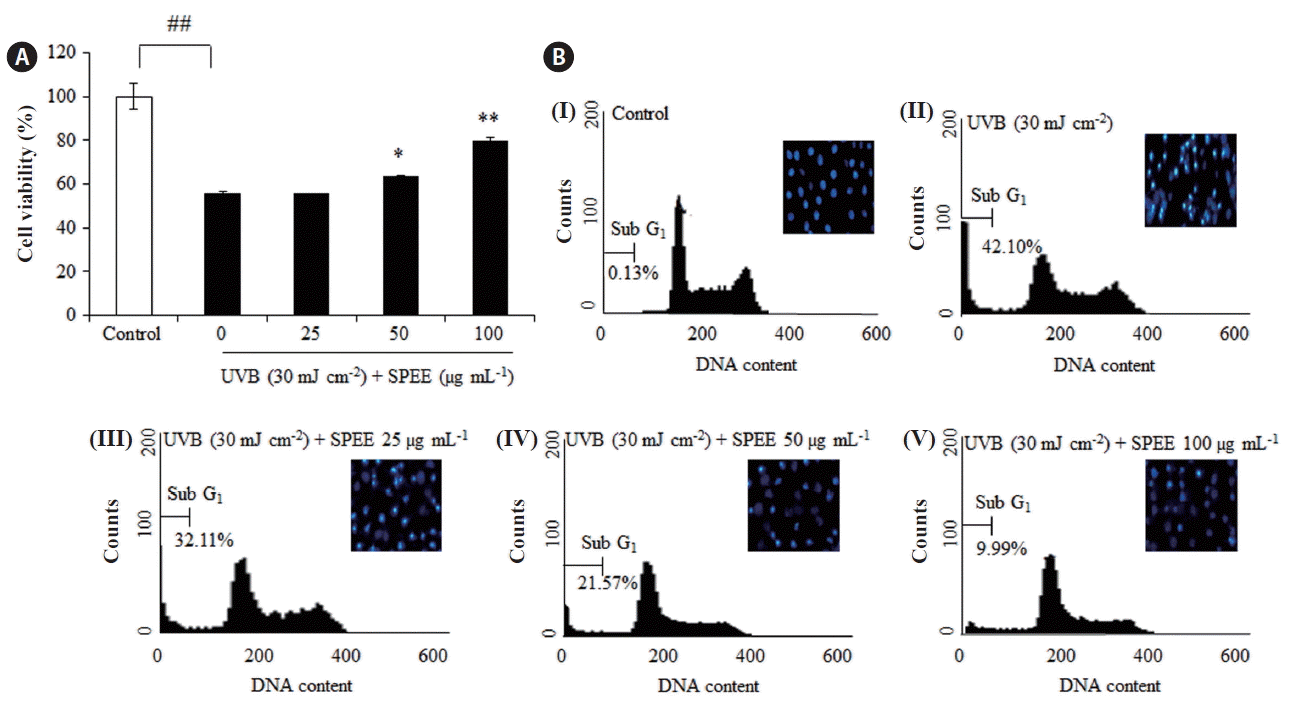
Protective effects of SPEE against UVB-induced apoptosis in HaCaT cells
The protective effects of SPEE on the damaged HaCaT cells induced by UVB irradiation were examined by MTT assay. Cells were treated with SPEE (25–100 μg mL−1) and exposed to 30 mJ cm−2 of UVB. Cells that were exposed to UVB showed 44.64% less viability compared with non-UVB irradiated cells (Fig. 3A). Treatment of cells with increasing concentrations (25, 50, and 100 μg mL−1) of SPEE prior to UVB exposure, enhanced cell viability by respectively 0.32, 8.22, and 23.33%. To assess the effect of SPEE on UVB-induced apoptosis of HaCaT cells, the amount of sub-G1 cells and apoptotic cells were analyzed using flow cytometry. As shown in Fig. 3B, treatment of HaCaT cells with SPEE (25, 50, and 100 mg mL−1) resulted in a decrease in the accumulation of cells in the sub-G1 phase by 9.99, 20.53, and 32.11%, respectively. Furthermore, direct observation by Hoechst staining revealed that nuclei with chromatin condensation, apoptotic bodies and widespread membrane blebbing were formed in cells that were exposed to UVB (Fig. 3B, cell images) and this was attenuated with SPEE treatment in a concentration dependent manner. This data suggested that SPEE treatment reduced UVB-induced apoptosis of HaCaT cells.
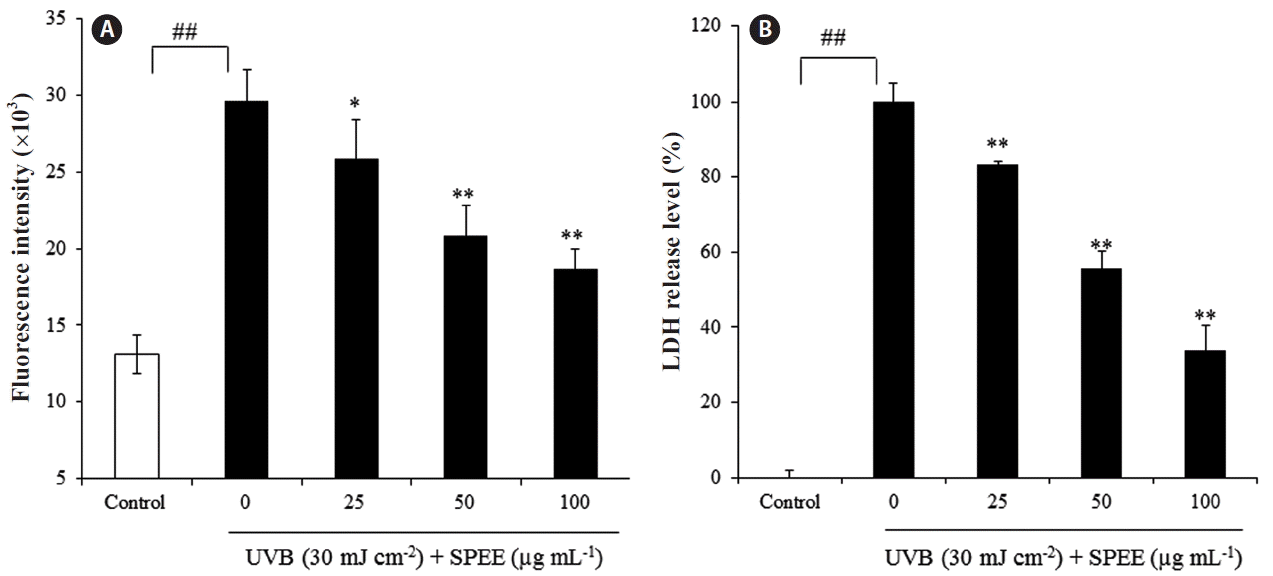
SPEE reduces UVB-induced cellular ROS generation and LDH release in HaCaT cells
To assess the effect of SPEE on the oxidative stress induced by 30 mJ cm−2 of UVB in HaCaT cells, intracellular ROS levels were determined by measuring the intracellular oxidation of 2,7-dichlorodihydrofluorescein (DCFH2). Exposure of DCFH2 loaded HaCaT cells to UVB, induced oxidative stress as assessed by DCF formation indicating the production of ROS (Fig. 4A). Treatment of cells with increasing concentrations of SPEE, protected cells from UVB-induced oxidative stress in a concentration-dependent manner.
LDH release as an indicative of cellular damage was also assessed. Cells exposed to UVB showed a significant increase in the level of LDH release, compared to non-UVB-exposed cells (p < 0.01) (Fig. 4B). The increased LDH release levels in UVB-exposed cells were reduced by 35.98% with 100 μg mL−1 of SPEE treatment (p < 0.01) (Fig. 4B).
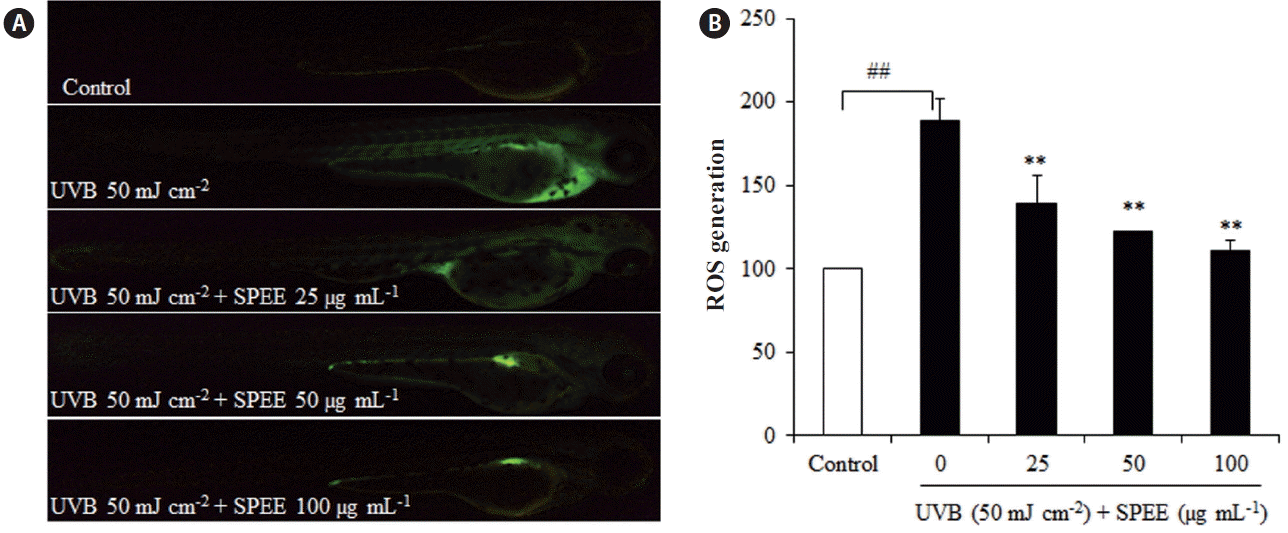
SPEE reduces ROS generation induced by UVB irradiation in zebrafish
The protective effect of SPEE on the damage induced by UVB was further assessed in in vivo model of zebrafish. Fish were treated with SPEE (25–100 μg mL−1) and exposed to 50 mJ cm−2 of UVB. Fig. 5A shows a typical fluorescence micrograph of a zebrafish. The control group, which was subjected to neither UVB nor SPEE, generated no fluorescence, whereas the group that was exposed to UVB, generated DCF fluorescence, suggesting an increase in the level of ROS by UVB irradiation in the zebrafish. However, when the zebrafish were treated with the different concentrations of SPEE prior to UVB exposure; a concentration-dependent reduction in the generation of ROS was observed (Fig. 5A & B). These results suggest a protective effect of SPEE against oxidative stress induced by UVB exposure in in vivo zebrafish model.
DISCUSSION
Exposure of cells to UVB radiation can result in an increased ROS generation, which is responsible for skin cancer development and photoaging (Raut et al. 2012). As ROS are implicated in skin damage by UVB, scavenging of these species could prevent the oxidative reactions and subsequently protect skin from the damaging effects of UVB (Masaki 2010, Kammeyer and Luiten 2015).
The phenolic phytochemicals from botanical antioxidants have been widely used for their ability to delay the process of photoaging by reducing the harmful effects of UV radiation through scavenging ROS (Heo et al. 2009, Ko et al. 2011, Ryu et al. 2015). Heo et al. (2010) reported diphlorethohydroxycarmalol isolated from Ishige okamurae showing strong protective effects against UVB-induced human fibroblast cell damage (Heo et al. 2010). Yang et al. evaluated the effects of quercetin against UVB-induced cell death in in vitro HaCaT cells and in vivo zebrafish model (Yang et al. 2012). This present work suggests the protective effects of SPEE against oxidative stress induced by UVB in in-vitro and in-vivo models.
Through serial fractionations of SPE, the gallic acid derivatives (SPEE) (Table 1) showing 774.42 ± 2.61 mg g−1 GAE was selected and analyzed in on-line HPLC-ABTS chromatogram at 680 nm, which is the specific wavelength for polyphenols reacting with the radical scavengers. Furthermore, the peaks derived from SPEE showing 70% or above of total antiradical activity were identified as gallic acid, ethyl gallate, and methyl gallate (Fig. 2B).
We evaluated the protective activity of SPEE on UVB-induced damage in HaCaT cells. The results in the study showed a protective effect of SPEE against reduction in cell viability caused by UVB irradiation. This observation was further studied for the protective effect of SPEE on UVB-induced apoptosis using flow cytometer and propidium iodide staining. The UVB-induced accumulation of cells in the sub-G1 phase was significantly decreased with SPEE pre-treatment in HaCaT cells, suggesting the protective effects of SPEE against sub-G1 cell cycle arrest, and apoptosis.
UVB-mediated apoptosis was also associated with an increase in the intracellular ROS levels, suggesting an activation of ROS-producing enzymes or impairment of the antioxidant pool which may predispose cells to lower defense against oxidative stress (Masaki 2010, Raut et al. 2012). To evaluate the activity of SPEE on the UVB-induced oxidative stress in HaCaT cells, we assessed the levels of ROS and LDH release with or without SPEE treatment. Treatment with SPEE prior to UVB exposure resulted in considerable protection of HaCaT cells against ROS generation and LDH release. We further examined the effect of SPEE against oxidative stress in in vivo zebrafish model. UVB exposure induced ROS generation in zebrafish, which was inhibited by treatment with SPEE. Collectively, these results suggested that SPEE reduced apoptosis induced by UVB via inhibiting ROS levels in HaCaT cells and zebrafish.
In conclusion, four main objectives in this study were achieved: (1) enhancement of phenolic content of Spirogyra sp. by serial fractionation, (2) determination of the radical scavenging activity of gallic acid derivatives of Spirogyra sp. (SPEE) based on on-line HPLC-ABTS, (3) separation of gallic acid, methyl gallate, and ethyl gallate from SPEE in CPC chromatogram. Among these compounds, ethyl gallate is isolated from Spirogyra sp. for the first time, and (4) examination of the protective effect of SPEE treatment prior to UVB exposure in in vitro HaCaT cells and in vivo zebrafish. This study may provide insight into the potential use of gallic acid derivatives from Spirogyra sp. for the treatment of skin damage caused by UVB.
SUPPLEMENTARY MATERIAL
Supplementary Table S1
The nuclear magnetic resonance spectroscopic data of ethyl gallate (http://www.e-algae.org).