Effect of elevated pCO2 on thermal performance of Chattonella marina and Chattonella ovata (Raphidophyceae)
Article information
Abstract
Ocean acidification and warming, identified as environmental concerns likely to be affected by climate change, are crucial determinants of algal growth. The ichthyotoxic raphidophytes Chattonella species are responsible for huge economic losses and environmental impact worldwide. In this study, we investigated the impact of CO2 on the thermal performance curves (TPCs) of Chattonella marina and Chattonella ovata grown under temperatures ranging from 13 to 34°C under ambient pCO2 (350 μatm) and elevated pCO2 (950 μatm). TPCs were comparable between the species or even between pCO2 levels. With the exception of the critical thermal minimum (CTmin) for C. ovata, CTmin for C. marina and the thermal optimum (Topt) and critical thermal maximum (CTmax) for both species did not change with elevation of pCO2 levels. While CO2 enrichment increased the maximum photosynthetic rates (Pmax) up to 125% at the Topt of 30°C, specific growth rates were not significantly different under elevated pCO2 for the two species. Overall, C. ovata is likely to benefit from climate change, potentially widening its range of thermal tolerance limit in highly acidic waters and contributing to prolonged phenology of future phytoplankton assemblages in coastal waters.
INTRODUCTION
Climate change scenarios have predicted a global mean temperature rise of 2.6–4.8°C by the end of this century (Core Writing Team et al. 2014), which could affect the changes in the biogeographic range of marine species and cause a substantial pole-ward extension of biodiversity across the North Pacific (Hazen et al. 2013). Concurrently, the global average sea surface temperature (SST) has increased by 0.7°C while decreasing the pH by 0.1 units since pre-industrial times. SST is expected to rise by a further 1.2–3.2°C, and pH is expected to fall by a further 0.3–0.4 units by the year 2100 according to the Representative Concentration Pathway 8.5 (RCP8.5) (Gattuso et al. 2015). These environmental changes could affect the potentially significant role of phytoplankton in removing CO2 from the sunlit ocean, as well as their growth and metabolism directly or indirectly in numerous ways (Raven 2017, Raven et al. 2020). Many studies have previously reported the response of marine algal growth to temperature warming and elevated CO2 (Kremp et al. 2012, Tatters et al. 2013, Brandenburg et al. 2019, Seto et al. 2019). In general, global warming benefits algal growth if the increased temperature is below the optimal temperature; beyond this temperature, it exerts adverse effects (Daufresne et al. 2009, Singh and Singh 2015). The fertilization effect of rising dissolved CO2 concentrations in surface waters may potentially contribute to the increase in growth and production, and range expansion of many harmful algal bloom (HAB) species (Fu et al. 2012, Flynn et al. 2015, Raven 2017, Raven et al. 2020). Additional CO2 is broadly beneficial for algal growth if nutrient availability is sufficient (Tatters et al. 2013, Thomas et al. 2017). Based on these findings, the effect of CO2 on algal photosynthesis and growth might be more noticeable at suboptimal temperatures than at supraoptimal temperatures. However, the optimal temperature for growth will depend on the levels of dissolved inorganic carbon (DIC), as well as on numerous physiological and ecological factors (Thomas et al. 2012, Kibler et al. 2015, Brandenburg et al. 2019).
In temperate and subtropical-tropical regions of the world, the fish-killing (ichthyotoxic) raphidophytes Cha-ttonella species have frequently caused serious damage to fisheries and mariculture industries in Korea (Kim et al. 2007), Japan (Imai and Yamaguchi 2012, Yamaguchi et al. 2018), China (Wang et al. 2006, 2017), North America (Lewitus et al. 2008, García-Mendoza et al. 2018), and Europe (Stacca et al. 2016, Satta et al. 2017, Zingone et al. 2020). The production of superoxide is associated with the growth of Chattonella, which is a strong candidate for the cause of its toxicity in fishes (Shikata et al. 2019). They inhabit the warm (~30°C) tropical waters around the equator (e.g., Chattonella subsalsa), extending to the colder (~5°C) waters of the Dutch Wadden Sea (e.g., Chattonella marina and Chattonella antiqua) (Vrieling et al. 1995). Chattonella species are often characterized by high phenotypic plasticity (Vidyarathna et al. 2020), which likely enables them to successfully colonize a wide range of environments. However, practical identification of Chattonella species is difficult because its pleomorphic cells and fragile nature (Coyne et al. 2005). A great morphological variability was observed as a function of laboratory culture conditions, as well as local environmental conditions; hence, there are no distinct morphological characteristics to discriminate these species (Demura et al. 2009).
Blooms of Chattonella tend to develop immediately after an increase in nutrient input during sunny weather and accelerate through their diel vertical migration behavior, in which cells move to the surface during the day to get access to light for photosynthesis and migrate toward the bottom during the night to have more access to nutrients throughout a diurnal cycle (Watanabe et al. 1991, Tilney et al. 2015, Shikata et al. 2019, Qiu et al. 2020). Given the potential role of resting cysts in bloom initiation of Chattonella species, the abundance of vegetative cells in the water column is not always relied on that of viable cysts in the sediments (Onitsuka et al. 2020). Meanwhile, the genus Chattonella was thought to be exclusively photosynthetic protists. However, the occurrence of mixotrophy has been described for C. marina and Chattonella ovata, which can feed on heterotrophic and autotrophic bacteria (Jeong et al. 2010, Jeong 2011). This may confer a competitive advantage to these species during nutrient shortage and add complexity to marine microbial food webs.
Chattonella species can grow over a wide range of temperatures. The appropriate temperature range for the growth of C. marina was from 15 to 30°C, with an optimal growth temperature of approximately 25°C (Imai and Yamaguchi 2012). The vegetative cells of C. marina were observed in Osaka Bay at temperatures ranging from 13 to 31°C and in the Seto Inland Sea, Japan from 18.8 to 28.0°C (Yamochi 1984, Yoshimatsu and Ono 1986). Moreover, an Australian strain of C. marina was able to grow at 10°C (Marshall and Hallegraeff 1999). The range of temperature have been experimentally determined for three strains of C. ovata isolated from Hiroshima Bay, and were found to be between 15 and 32.5°C (Yamaguchi et al. 2010). In the Seto Inland Sea, the vegetative cells of C. ovata found at a wide range of temperatures (15.8–31.3°C), showing the highest abundance at temperatures of 26.0–29.5°C in summer (Imai and Yamaguchi 2012). The optimal growth temperature for C. ovata varied between 25 and 30°C and was higher than that for C. antiqua, C. marina, and C. subsalsa, which showed an optimum growth temperature at 20–30°C, significantly reduced growth rates at 10–16°C, and no growth at 4°C (Kahn et al. 1998, Zhang et al. 2006, Imai and Yamaguchi 2012). The Korean strain of C. marina var. marina and C. marina var. ovata rarely grew at 10°C, and their optimal temperatures were 25 and 30°C, respectively (Noh 2009). In Korean waters, Chattonella species generally occur when water temperature ranges from 14.5 to 30.5°C and grow rapidly when temperature ranges from 23.1 to 30.5°C (Jeong et al. 2013). Based on these data, surface water temperature is often presumed to be a primary controlling factor for the Chattonella red tide in Korean waters.
The effects of temperature on fitness-related traits (i.e., photosynthesis and growth) within the survival range have been experimentally determined for a wide variety of algal species and strains (Hallegraeff 2010, Thomas et al. 2012, Kontopoulos et al. 2020). The relationship between trait rate and temperature can be visualized as a thermal performance curve (TPC). Two features of a TPC common to all ectotherms are unimodality and negative skewness (Angilletta 2006, Kingsolver 2009). The rate values increase with temperature until a critical point or optimal temperature (Topt), after which they drop rapidly. The range of temperatures between maximum temperature or critical thermal maximum (CTmax) and the minimum temperature or critical thermal minimum (CTmin) is defined as the thermal niche or thermal tolerance (Ttol), and can vary according to an organism’s phenotypic plasticity, physiological traits, and evolutionary history (Thomas et al. 2016, Jin and Agustí 2018, Vidyarathna et al. 2020). To understand the capacity for adaptation of TPC in different thermal environments, it is important to investigate how the shape of the TPC changes across species. It is also necessary to evaluate how the TPC of growth rate is affected by rising CO2 concentration in a globally important HAB species, including Chattonella. However, limited information is available regarding the physiological constraints that set thermal tolerance extents in Chattonella species and how adaptation can overcome these constraints to enhance thermal performance. Nevertheless, this research will be essential when trying to predict the production and global distributions of Chattonella in the face of climate change.
The aims of this study were to determine the cardinal temperatures for the growth rate of C. marina and C. ovata under ambient and elevated CO2 conditions from TPCs and to subsequently assess the effect of CO2 on their photosynthesis and growth responses at the optimal growth temperature. To this end, C. marina and C. ovata were grown under two pCO2 levels (350 and 950 μatm), in combination with five temperature (13, 20, 26, 30, and 34°C) treatments, and the interactive effects on growth rate and maximum photochemical efficiency were assessed in tightly controlled laboratory experiments.
MATERIALS AND METHODS
Cultures
The strains of two raphidophytes, Chattonella marina (LIMS-PS-2900) isolated from Jangmok in January 2007 and C. ovata (COKP9909) isolated from Gyeokpo in September 1999, were obtained from the Culture Collection of the Korea Institute of Ocean Science & Technology (KIOST) and the cultures were maintained by Prof. HJ Jeong at Seoul National University, respectively. Culture strains were grown and maintained in 250-mL Erlenmeyer flasks containing 200 mL of modified f/2 medium lacking silica at 20–23°C. The medium was prepared in sterilized natural seawater, with a salinity of 32–34 at pH 8.0. A 12 : 12 h light-dark cycle with 120–150 μmol photons m−2 s−1 light intensity was provided by 36 W daylight fluorescent lamps (Dulux L 36W/865; Osram, Münich, Germany). Cultures were transferred weekly to keep them in exponential growth phase until the start of the experiment.
Experimental setup
C. marina and C. ovata were semi-continuously cultured for 7 days after a 3-day batch culture under each treatment condition (see below). During the semi-continuous culture, Erlenmeyer flasks (100 mL capacity) containing 100 mL of cultures were kept at the lower section of the exponential growth phase (Supplementary Fig. S1) and were optically thin to avoid self-shading, nutrient limitation, and to minimize CO2 drift. The culture flasks were shaken gently twice a day to ensure the mixing of cell suspensions. The five temperature levels (13, 18, 25, 30, and 34°C) were combined with two CO2 partial pressures (pCO2); 350 μatm (ambient pCO2 treatment) or 950 μatm (elevated pCO2 or ocean acidification [OA] treatment), and experiments were performed in triplicate. These OA mimicking values were based on the IPCC WG RCP 8.5 scenario. Every 24 h, the cultures were diluted with the respective f/2 media that had been adjusted to the two target pCO2 levels, to a starting cell concentration of 100 cells mL−1 in C. marina and 400 cells mL−1 in C. ovata. To attain and maintain the target pCO2 or pH values, the culture was additionally adjusted by adding an appropriate amount of CO2-saturated seawater. No more than 10 μL of CO2-saturated seawater was used. CO2-saturated seawater was prepared by bubbling reagent-grade CO2 through seawater within gas tight containers, which caused the medium pH to drop below 5. Prior to every dilution, 5 mL of culture were collected in triplicate from semi-continuous cultures to measure photosynthetic efficiency, spectrophotometric pH (total scale), and cell counts.
Temperature gradients
The five target temperatures, ranging from 13 to 34°C were obtained using a plexiglass aquarium (45 cm × 20 cm × 20 cm) with a thermostatically controlled water bath, filled with deionized water and equipped with a heater and a cooler, which allowed control of the temperature with a precision of ±0.3°C (Supplementary Fig. S1). The temperature inside the aquarium was logged every 30 min using a HOBO temperature logging device (Onset Computer Corporation, Pocasset, MA, USA). Erlenmeyer flasks (100 mL capacity) containing 100 mL of culture were placed in a plexiglass aquarium in three replicates, under 120 μmol photons m−2 s−1 of photosynthetically active radiation (400–700 nm) with a 14 : 10 h light-dark cycle.
Inorganic carbon chemistry
The inorganic carbon chemistry was determined using the CO2SYS program (Lewis and Wallace 1998) using total alkalinity (AT), total pH scale, salinity, and temperature of culture seawater. The AT was measured using potentiometric acid titration in a semi-closed cell system, consisting of a Metrohm 765 Dosimat titrator (Metrohm, Zofingen, Switzerland) connected to a pH meter (Orion 920A; Thermo Fisher Scientific, Waltham, MA, USA). The added volume of acid (0.13 N HCl) and electromotive force were recorded using Q-basic software during each titration (Millero et al. 1993). The pH of the culture samples was determined using a high-resolution spectrophotometer (Agilent 8453, UV-VIS Spectrophotometer; Agilent Technologies, Palo Alto, CA, USA). Absorbance at wavelengths of 434, 578, and 730 nm of seawater samples were measured at 25°C with m-cresol purple indicator dye (Sigma-Aldrich Chemical Co., St. Louis, MO, USA) (Dickson 1993). The method employed was essentially the same as that described by Kim et al. (2018).
Maximum photochemical efficiency of photosystem II
The photosynthetic activity of Chattonella cultures was determined using pulse amplitude-modulated (PAM) chlorophyll-a fluorometry (Phyto-PAM fluorometer; Walz GmbH, Effeltrich, Germany). The maximum efficiency of photosystem II (PSII) photochemistry (Fv/Fm) was measured for each culture after being acclimatized to darkness for 10 min, where Fv (variable fluorescence) is calculated as the difference between Fm and Fo fluorescence, Fo is the fluorescence level in the absence of actinic (photosynthetic) light for open PSII reaction centers, and Fm is the maximum fluorescence at closed reaction centers induced by a short pulse of saturating light (Genty et al. 1989).
Photosynthesis vs. irradiance curves (P-E curves)
Photosynthetic oxygen evolution was measured by inserting a Clark-type O2 microelectrode with a tip diameter of 500 μm (OX-MR; Unisense, Aarhus, Denmark) connected to a pico-amperemeter (PA2000; Unisense) into a 4 mL micro-respiration chamber (Unisense) in a thermostatically controlled water bath at 30°C. The O2 concentration of cultures was recorded every minute for 15–20 min at all light intensities. The oxygen electrode was two-point calibrated by measuring between 0 and 100% saturated seawater. A 250 W halogen lamp (KL 2500 LCD; Schott, Mainz, Germany) illuminated the setup, and variable levels of irradiance were obtained by using shade screens with different densities (0, 21, 42, 80, 171, 326, and 652 μmol photons m−2 s−1). Real-time oxygen measurements were initiated by measuring respiration in darkness. Photosynthesis was subsequently measured at increasing levels of irradiance. Rates of O2 consumption or release were calculated from incubation periods with constant changes in O2 concentration over a minimum of 10 min. A full photosynthesis-irradiance (P-E) curve for each sample lasted for 2–3 h, and three replicate P-E curves were run. Photosynthetic rates were expressed in units of μmol O2 cell−1 d−1. The photosynthetic parameters of the P-E curves were estimated by the non-linear regression model of Platt et al. (1980) to identify photosynthetic traits. The maximum photosynthetic rate in the absence of photoinhibition (Pmax, μmol O2 cell−1 h−1), photosynthetic efficiency under non-saturating irradiance or the initial slope of the P-E curve (α, μmol O2 cell−1 h−1 [μmol photons m−2 s−1]−1), and irradiance at onset of saturation for P-I curves or light-saturation parameter (Ek, μmol photons m−2 s−1) were determined using the least squares curve fitting technique included with the software Grapher ver. 12 (Golden Software Inc., Golden, CO, USA).
Growth
The number of live cell was counted daily in a 1 mL Sedgewick-Rafter counting chamber under a compound microscope. If there was not enough time for this task, samples were preserved with 5% Lugol’s iodine solution for later enumeration. Growth was calculated as the specific growth rate (μ d−1) as follows:
, where N0 and Nt denote the cell concentration at the initial and after a time interval of the experiments, and Δt is the corresponding incubation time (d) of exponential growth (Supplementary Figs S2 & S3).
Cardinal temperature estimation
The growth-temperature (μ-T) curves were described by fitting the cardinal temperature model with inflection (CTMI) described by Rosso et al. (1993):
for Tmin ≤ T ≤ Tmax, where T is the temperature (°C), μ is the specific growth rate on d−1, Topt is the optimal temperature at which the specific growth rate is maximal, and μopt is the specific growth rate at Topt. CTmin and CTmax are the hypothetical lower and upper critical temperature (critical thermal minimum and maximum), respectively, through which the specific growth rate is zero. Because an estimation of CTmax and CTmax at zero has limited biological meaning, we calculated CTmin and CTmax as the temperatures at which performance was 5% of the maximum performance. The range between CTmin and CTmax is termed the tolerance range (Ttol). The thermal performance breadth (TB80) was also determined and was defined as the temperature range through which the growth rate was close to optimal (defined as the 80th percentile of the CTMI fits).
Statistical analyses
The main and combined effects of two pCO2 (ambient and elevated) and five temperature treatments with respect to all experimental parameters (carbonate chemistry, growth rate, and maximum efficiency of PSII photochemistry) were evaluated by two-way analyses of variance (ANOVAs). Prior to the statistical analysis, data sets were tested for conformity to normality and for homogeneity of variances using the Shapiro-Wilk and Levene’s test, respectively. Two-way ANOVA was also performed to determine the effects of pCO2 on species and their interaction based on the photosynthetic and growth rates at the optimal growth temperature (30°C). If a significant ANOVA was found, differences among the group means were tested using Scheffé’s post hoc tests for multiple comparisons. All statistical analyses were performed using SPSS Statistics ver. 25 (IBM Corp., Armonk, NY, USA).
RESULTS
The two target pCO2 levels 350 and 950 μatm were successfully maintained at all temperature treatments throughout the experimental period (Table 1). Regardless of temperature, the pCO2 (mean ± standard deviation) in cultures of C. marina was significantly higher under elevated pCO2 (966.7 ± 164.6, n = 300) than under ambient pCO2 (413.5 ± 112.6, n = 366) (Scheffé’s test after ANOVA; p < 0.001). The pCO2 of C. ovata cultures was also significantly higher under elevated pCO2 (946.2 ± 130.5, n = 192) than under ambient pCO2 conditions (302.7 ± 61.6, n = 189) (Scheffé’s test after ANOVA; p < 0.001) at all five temperature treatments. Similar trends for the pCO2 were apparent for the pH values, which showed a range of 7.80 to 8.24 and 7.59 to 7.91 in cultures of C. marina of ambient and elevated pCO2 levels, respectively. The pH in cultures of C. ovata was also significantly lower under elevated than under ambient pCO2 levels (Scheffé’s test after ANOVA; p < 0.001), with ranges between 7.93 to 8.27 and 7.56 to 7.87, respectively (Table 1). In addition, the concentrations of bicarbonate (HCO3−) increased significantly with decreasing pH, whereas carbonate (CO32−) decreased under elevated pCO2 conditions. On the initial and final day of the experiment, total alkalinity (AT, μmol kg−1) remained constant with average and standard deviation values of 2,331.2 ± 15.3 (n = 6) and 2,318.2 ± 11.17 (n = 30) in the cultures of C. marina, and 2,260.6 ± 5.0 (n = 6) and 2,273.2 ± 6.6 (n = 30) in the cultures of C. ovata, respectively.

A summary of the seawater carbon chemistry of the Chattonella marina and C. ovata cultures at two target pCO2 (350 and 950 μatm) and five temperatures (13, 20, 26, 30, and 34°C) during the experiment
The specific growth rate (μ) was similar between C. marina and C. ovata, both at the ambient and elevated pCO2 (Fig. 1, upper panel). They grew over a wide range of temperatures (13–34°C), with optimum growth temperatures around 30°C under both pCO2 treatments. Cultures grown under elevated pCO2 exhibited an average increase of 6% in growth rates of C. ovata only, compared with those grown under ambient pCO2 across temperature treatments. Cultures of C. marina grown at 13°C showed the lowest growth rates, whereas cultures maintained at 34°C had the highest growth rate. The growth rate of increase in C. ovata with temperature increased to 30°C and then decreased abruptly at 34°C, maintaining only 10–15% of the maximum growth rate. ANOVA demonstrated that the specific growth rates of C. marina and C. ovata grow under two pCO2 levels over a range of temperatures, were significantly affected by temperature (df = 4, F = 77.3, p < 0.001 for C. marina; df = 4, F = 110.2, p < 0.001 for C. ovata), but not by either pCO2 treatments or pCO2 and temperature interaction. Scheffé’s post hoc analysis revealed significant differences between the growth rate at the lowest temperature (13°C) and those at the other temperatures for C. marina (p < 0.001). There were also significant differences in growth rates of C. ovata at 13 and 34°C compared with those at the rest of the temperatures examined (p < 0.001). Noticeably, the growth rates of C. ovata in the range of temperatures below the optimal were slightly higher at elevated pCO2 than ambient pCO2 treatments.
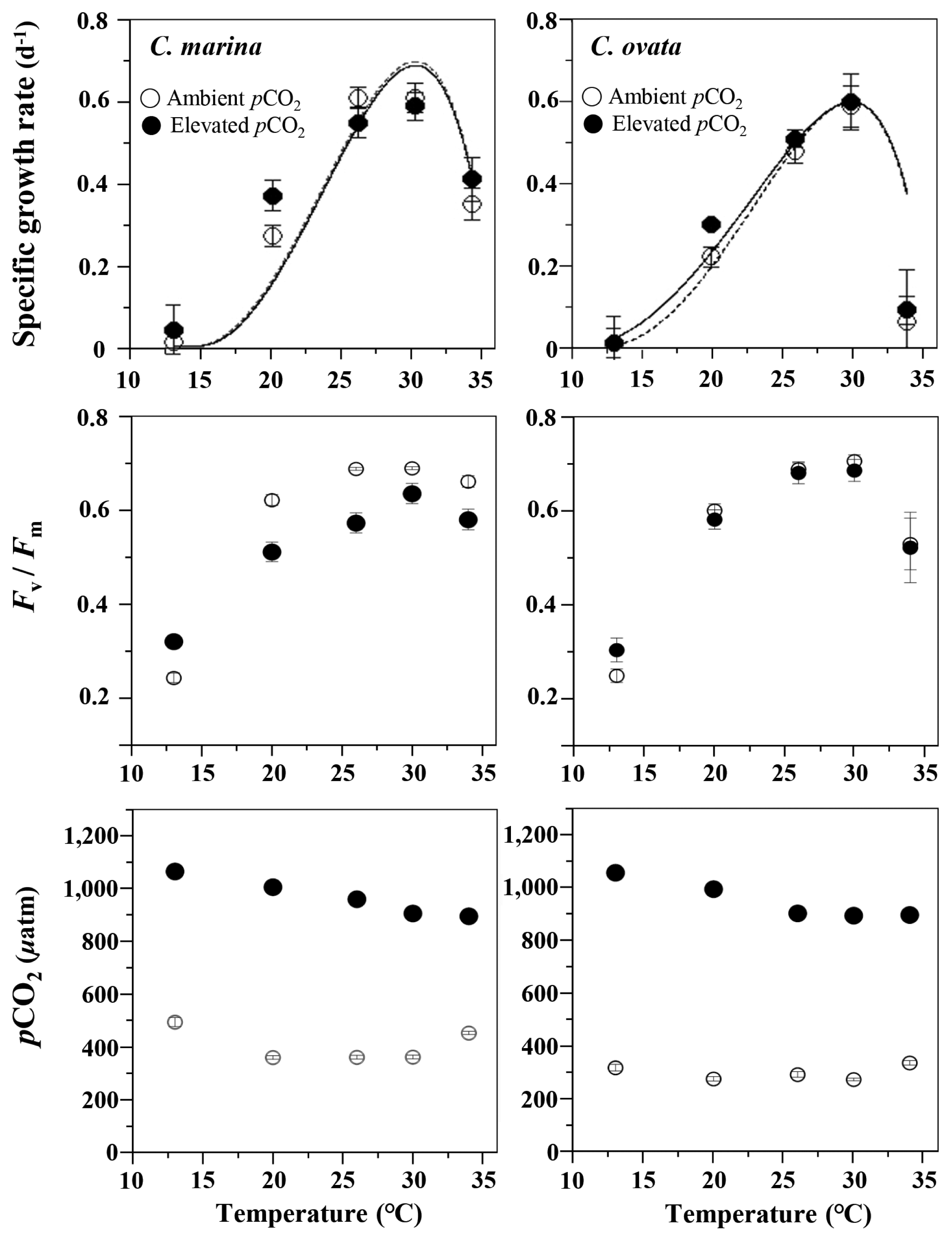
Specific growth rate (μ d−1) and maximum photochemical efficiency (Fv/Fm) of Chattonella marina and C. ovata in response to pCO2 and water temperature treatments. Vertical bars indicate the mean ± standard error (n = 3) for ambient (open circles) and elevated (filled circles) pCO2 treatments. Lines represent the best fit of the data to the Cardinal Temperature Model with Inflection (CTMI) model of Rosso et al. (1993) with r2 of 0.959 (open circles) and 0.918 (filled circles) in C. marina, and 0.876 (open circles) and 0.805 (filed circles) in C. ovata.
Unlike growth rate, the maximum photochemical efficiency (Fv/Fm) of C. marina and C. ovata increased under both pCO2 levels as temperatures increased from 13 to 30°C, showing the capacity to maintain photosynthesis at 34°C (Fig. 1, middle panel). The Fv/Fm was greatest at the optimal growth temperature under both pCO2 conditions, and exhibited similar trends in both species, with cultures receiving the 20, 26, 30, and 34°C treatments, producing 0.5–0.7, except at 13°C (Fig. 1, middle panel, Supplementary Fig. S4). Both temperature and pCO2 treatments had significant effect on the Fv/Fm of C. marina cultures (df = 4, F = 234.4, p < 0.001 for temperature; df = 1, F = 39.0, p < 0.001 for pCO2). In addition, an interaction effect of temperature and pCO2 on the Fv/Fm of C. marina was observed (df = 4, F = 15.2, p < 0.001). For C. ovata, only the temperature significantly influenced the Fv/Fm (df = 3, F = 259.8, p < 0.001). In both pCO2 conditions, Scheffé’s post hoc tests showed that Fv/Fm value at 13°C was significantly lower (p < 0.001) compared to that at the other temperatures.
Application of CTMI model to the growth rate resulted in cardinal temperatures, that is critical thermal minimum (CTmin), critical thermal maximum (CTmax), thermal optimum or optimal temperature (Topt). Under ambient pCO2 conditions, the CTmin, CTmax, and Topt for the growth rate of C. marina were estimated to be 17.0, 35.6, and 30.0°C, respectively. Under elevated pCO2 conditions, the CTmin, CTmax, and Topt were estimated to be 17.1, 35.7, and 30.0°C, respectively (Table 2). Using the CTMI model, the CTmin, CTmax, and Topt for the growth rate of C. ovata under ambient pCO2 conditions were estimated to be 15.1, 35.7, and 30.0°C, respectively. Under elevated pCO2 conditions, a CTmin of 13.6, CTmax of 35.7, and Topt of 30.0°C were obtained (Table 2). Overall, there was only a slight decrease in the CTmin in response to elevated pCO2, but no change in either CTmax and Topt between the ambient and elevated pCO2 treatments.

Parameters of thermal performance curve for the growth rate of Chattonella marina and C. ovata in response to pCO2 (ambient 350 and elevated 950 μatm), as calculated by the CTMI (Cardinal Temperature Model with Inflection) model
A more comprehensive assessment of the light-dependence of photosynthesis or oxygen evolution rate was performed at the optimal growth temperature of 30°C (Fig. 2). The photosynthetic efficiency under non-saturating irradiance or initial slope (α) of the P-E curve showed little response to pCO2, but the maximum photosynthetic rate (Pmax) increased by 78% from 0.106 μmol O2 cell−1 h−1 at ambient conditions to 0.189 μmol O2 cell−1 h−1 at elevated pCO2 levels for C. marina. A corresponding increase by 125%, from 0.039 to 0.088 μmol O2 cell−1 h−1, was seen in C. ovata. The photosynthetic photon flux density at which the highest photosynthetic rate (Ek) occurred also increased with pCO2 levels from 96 μmol photons m−2 s−1 at ambient pCO2 to 171 μmol photons m−2 s−1 at the elevated pCO2 for C. marina. The light-saturation parameter (Ek) for C. ovata showed a similar response to pCO2, but was much lower than that of C. marina.
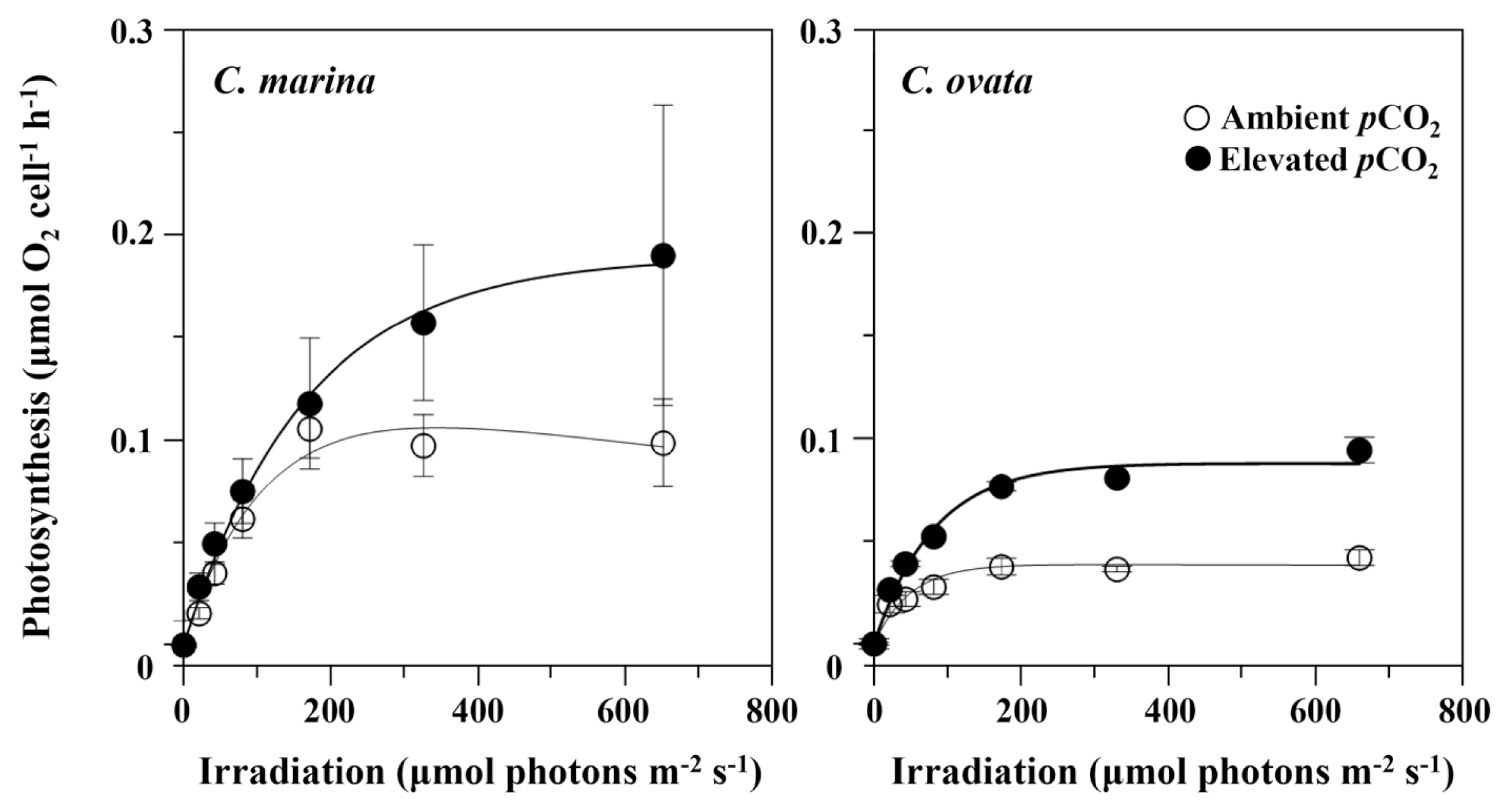
Effect of elevated pCO2 on gross photosynthesis or oxygen evolution rates of Chattonella marina and C. ovata in response to photosynthetic photon flux density at optimal growth temperature (30°C). Vertical bars indicate the mean ± standard error (n = 3) for ambient (open circles) and elevated (filled circles) pCO2 levels. Lines represent the best fit of the data to the photoinhibition model of Platt et al. (1980).
All measured growth rates at the optimal growth temperature (30°C) were generally similar between the CO2 treatments (Fig. 3). Analysis of the growth rate as a function of pCO2 indicated that it was not changed by increasing CO2, and only a small difference was found between the two species (Fig. 3). For C. marina, the mean growth rates at ambient and elevated pCO2 were 0.44 ± 0.03 and 0.47 ± 0.03, respectively, while the comparable values for C. ovata were 0.38 ± 0.05 and 0.48 ± 0.04, respectively. The two species did not differ significantly in response of specific growth rate to pCO2 (df = 1, F = 0.351, p = 0.578). The growth rates of both species exposed to elevated pCO2 were slightly greater than those exposed to ambient conditions, but the difference was not significant (df = 1, F = 2.379, p = 0.133). In addition, the interaction between the CO2 concentration and the species was not statistically significant (df = 1, F = 0.871, p = 0.358).
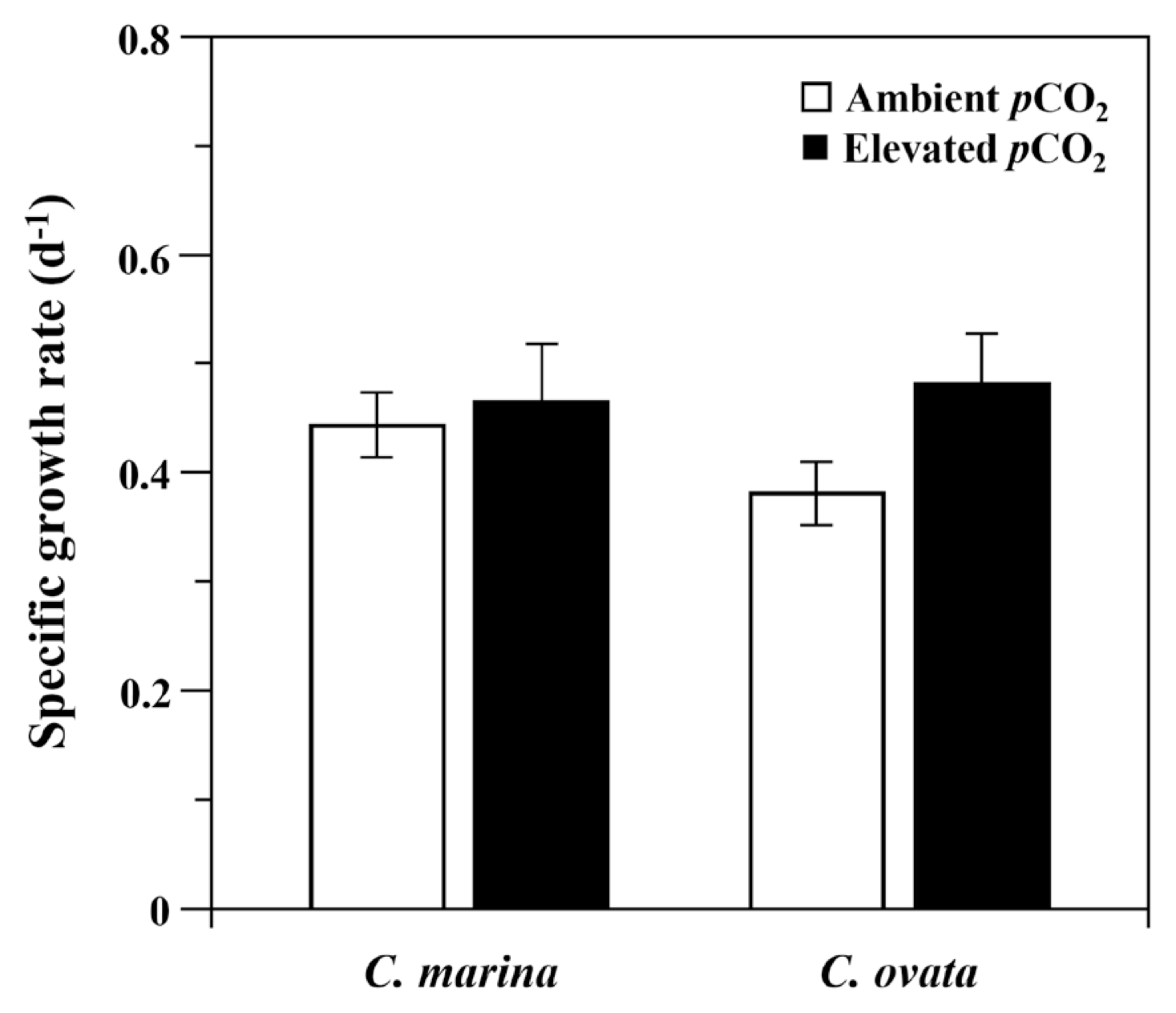
Specific growth rate (μ d−1) of Chattonella marina and C. ovata at optimal growth temperature (30°C). Vertical bars indicate the mean ± standard error (n = 3) for ambient (open column) and elevated (filled column) pCO2 levels. None of the differences were statistically significant by two-way ANOVA (p > 0.001).
Our key findings for Chattonella were as follows: (1) the maximum performance or maximum growth rate (μmax) was slightly higher in C. marina than in C. ovata; (2) thermal tolerance (Ttol) and thermal performance breadth (TB80) were slightly greater in C. ovata than in C. marina; (3) CO2 affected the Ttol and TB80 in C. ovata but not in C. marina; (4) at the optimal temperature (Topt), CO2 affected the maximum photosynthetic rate (Pmax), but there was no effect of CO2 on the specific growth rate (μ) in both species.
DISCUSSION
Chattonella marina var. ovata strains from Korea rarely grew at 10°C, and slightly at 15°C, with growth increasing to a maximum of 0.62 d−1 at 30°C (Noh 2009). C. marina var. marina showed a pattern similar to that of C. marina var. ovata, although its peak growth rate was 0.64 d−1 at 25°C (Noh 2009). In Korean waters, Chattonella species were identified when the water temperature was 14.5–30.5°C, and the water temperature during Chattonella red tides was 23.1–30.5°C (Jeong et al. 2013). These species were also found at 10–33°C in the waters of other countries and formed red tides at 24.0–31.0°C (see Jeong et al. 2013, table 4). Therefore, Chattonella occurrence and bloom dynamics are likely to respond similarly to water temperature in Korean waters as in the waters of other countries.
To better predict how species respond to changes in temperature, it is necessary to depict the shape of TPC (Kontopoulos et al. 2020). TPCs describe an organism’s eco-physiological trait according to temperature, which generally has a concave unimodal shape. The shape of TPC is similar across taxa, and much of the variation can be attributed to several factors, including genetics, seasonal, latitudinal, or acclimation capacity (Kingsolver 2009, Clusella-Trullas et al. 2011, Boyd et al. 2013, Bernhardt et al. 2018). In our results, all TPCs were non-symmetrical around the optimal temperature (30°C), exhibiting negative skewness. The curves exhibited a sudden sharp decline in growth rate accompanying small changes in temperature closer to the upper tolerance limit, and a smaller change in growth rate with changes in temperature near the lower limit (Fig. 1, upper panel). This indicates that when acclimated to Topt, fitness-related traits (i.e., photosynthesis and growth) are more significantly reduced by warming than cooling. Other research has suggested that OA has great potential to increase phytoplankton growth rates in regions of the ocean where temperature is equal to or less than the Topt (Boatman et al. 2017). Considering the SST anomaly from KNIFS (Korean National Institute of Fisheries Science) and the Topt of 30°C estimated for two Chattonella species in the present results, future warming is likely and will contribute to further intense, more frequent, and longer Chattonella red tides in Korean waters.
In a previous study, laboratory experiments and meta-analyses were conducted using TPCs to understand the differences in functional traits and physiological responses to temperature between phytoplankton species and between strains (Boyd et al. 2013). The CTMI model is derived from the prediction of a specific growth rate as a function of temperature in species or strains on the basis of empirical knowledge of cardinal temperatures and best visual fit, with respect to the available data (Rosso et al. 1995). In this study, the cardinal temperature parameters were calculated using the CTMI model for each Chattonella species and CO2 treatment to describe the growth rate change, including the biological significance obtained at different temperatures (Fig. 1, upper panel). Two other essential parameters derived from TPCs are thermal tolerance (Ttol = CTmax - CTmin) and thermal performance breadth (TB80). TB80 is the range of water temperatures over which the performance is greater than a given proportion (80%) of μmax (e.g., TB80 = 80% of μmax) (Angilletta 2006). Because cultures are likely to continue to be active at temperatures beyond Topt, TB80 represents the temperature range where cultures exhibit ≥80% of their maximal growth rate or performance, providing a conservative optimal performance range beyond which cultures begin to lose the ability to grow. Cardinal temperatures by CO2 interactions have been observed for only C. ovata, and there are slight increases in Ttol and TB80 to elevated CO2 without increasing growth rate (Table 2). Temperatures above TB80 result in steep decreases in performance until cultures become non-active, corresponding with the CTmax. The maximum temperature limit for growth or CTmax was similar between the two species and the two CO2 treatments. How future increases in surface temperature will influence the distribution will depend on the capacity for Chattonella species to adapt by increasing its CTmax or upper thermal tolerance limit. Interestingly, we found that C. marina and C. ovata have 5°C higher CTmax values than that did C. marina or C. Antiqua (Noh 2009, Imai and Yamaguchi 2012). There were no differences in CTmax and Topt between species or CO2 treatments, with values of 35.7 and 30.0°C, respectively (Table 2). However, Ttol and TB80 depend on species and CO2 treatments in C. ovata; specifically, the effect of the CO2 on growth of C. ovata was more pronounced at suboptimal than supraoptimal temperatures. Although no change in the CTmin of 17°C for C. marina, the CTmin for C. ovata growth was affected by CO2 and dropped from 15.1°C at ambient to 13.6°C at elevated CO2. The drop in CTmin with CO2 enrichment may even be underestimated, as it represents the temperature at which the growth rate exhibits ≤5% of the maximum performance. The thermal tolerance or niche showed a relatively wide temperature range from 17.0–35.7°C in C. marina and 13.6–35.7°C in C. ovata, is a key factor that sets the limits of their geographic distribution. Therefore, the parameters of TPC for growth rate extracted from each species-specific fit are necessary to better predict the impact of future climate change on large-scale biogeographic patterns in marine systems (Bestion et al. 2018, Kontopoulos et al. 2020).
Under light-saturated conditions, the maximum photochemical efficiency (Fv/Fm) peaked near Topt and remained high at temperatures above Topt, indicating that PSII was not damaged by high temperatures, irrespective of pCO2. The target sites of elevated temperature-induced or thermal damage in plants are the oxygen-evolving complex with the associated cofactors in PSII, the carboxylation activity of ribulose bisphosphate carboxylase-oxygenase (Rubisco), and the ATP generating system (Allakhverdiev et al. 2008). The photosynthetic rate of algae is limited by the activity of Rubisco, which in turn is influenced by various environmental factors, including light, temperature, nutrients, and DIC. Inorganic carbon acquisition strategies are likely to vary among species, due to differences in the operation of their carbon concentrating mechanisms (CCMs), which maintain elevated CO2 concentrations in the vicinity of Rubsco to ensure effective carboxylation and, consequently, efficient carbon fixation (Thoms et al. 2001, Giordano et al. 2005). The current CO2 concentrations are insufficient to saturate Rubisco carboxylation activity through passive CO2 diffusion into the cell in eukaryotic algae (Tortell et al. 2000, Beardall et al. 2009). Under such carbon-limited conditions, marine algae that utilize HCO3−, or rely on active CO2 uptake, could benefit from higher pCO2 by downregulating their CCMs, as such CCMs have high energy demands (Rost et al. 2008, Beardall et al. 2009, Eberlein et al. 2016, Van de Waal et al. 2019). Consequently, an increase in DIC availability in the water will result in a rise in photosynthetic rate and subsequent growth, in part because high CO2 concentrations inhibit the oxygenase reaction of Rubisco and reduce the loss of CO2 through photorespiration.
While the DIC was significantly higher in elevated pCO2 relative to the ambient pCO2 treatments, the carbonate chemistry in both species cultures significantly differed between the ambient and elevated pCO2 conditions (Table 1). Specifically, the pH decreased by 0.24–0.45 units relative to the ambient pH when the pCO2 level was elevated by appropriate injection of CO2-saturated seawater. As a consequence, the process made it possible to maintain carbonate chemistry throughout the entire experimental period. There are changes in carbonate system speciation; increasing pCO2 (which is equivalent to increasing DIC at constant AT) leads to an increase in H+ (decreasing pH) and bicarbonate (HCO3−) concentrations and decreasing carbonate (CO32−) concentrations (Table 1). There are two basic experimental approaches to adjust seawater CO2; either changing total alkalinity (AT) at constant DIC by NaOH and / or HCl addition or changing DIC at constant AT (e.g., aeration with air at target pCO2, injection of CO2-saturated seawater, combined NaHCO3 / Na2CO3 and HCl additions) (Gattuso et al. 2010). Experimentally, DIC or AT can be manipulated in different ways, depending on whether the initial seawater values are lower or higher than intended. While AT can be manipulated by adding strong acids or bases such as HCl or NaOH, various methods exist to increase or decrease pCO2 or DIC. In this study, the target pCO2 or DIC can be adjusted by injection of certain amounts of CO2-saturated seawater, which can easily be prepared by aeration with pure CO2 gas. Naturally occurring anthropogenic CO2 intrusion of the surface ocean increases seawater pCO2 and DIC without changing AT. Again, AT is not influenced by CO2 uptake and remains constant (Schulz et al. 2009). Furthermore, the unarmored and fragile cell nature of Chattonella are sensitive to the physical stresses, and the gas bubbles produced by aeration with air at the target pCO2 seem to be the cause of cell damage or breakage.
Despite different growth responses in different HAB species to high CO2, a number of HABs have generally been shown to experience increased growth rates under elevated CO2 concentrations; however, some other studies showed no increase in growth rate with pCO2 above the present air-equilibrium concentration. Even different isolates within a given species may have opposite responses to high and low CO2 for eukaryotic species (see Raven et al. 2020). It seems that different HABs differ in their capacity and strategy to adjust metabolism to high CO2 conditions. Furthermore, almost nothing is known about the effects of CO2 enrichment on photosynthesis and growth of the Raphidophycean flagellate Chattonella. In this study, the photosynthesis-irradiance (P-E) curves indicated that the maximum photosynthetic rate (Pmax) and light-saturation parameter (Ek) were significantly higher under elevated CO2 than under ambient CO2 conditions in both species (Fig. 2). The Pmax increased by about 78% for C. marina and 125% for C. ovata, as pCO2 increased from 350 to 950 μatm. However, there were no significant differences in growth rates between pCO2 levels or between species, although growth of both species seem to be higher under elevated pCO2 than under ambient pCO2 conditions (Fig. 3). These results appear to indicate that the cell growth and accumulation of Chattonella are not affected by CO2 enrichment, despite the increase in oxygen evolution rate being associated with high intercellular CO2 concentrations. Why does the maximum photosynthetic rate (Pmax) not result in increased growth of Chattonella under elevated CO2 levels? The mechanisms responsible for this discrepancy are not clear but may be related to the activation of CCM, which facilitate active transport of CO2 and / or HCO3− into the cytoplasm and chloroplast. With regard to the occurrence of a mechanism for energized transport of inorganic carbon, a number of CCM genes have been identified from de novo assembled transcriptomes in marine raphidophyte algae (Hennon et al. 2019). However, an essential feature of CCMs has yet to be confirmed with further experimental evidence of physiological mechanisms (Hennon and Dyhrman 2020, Raven et al. 2020). Some CCA genes, such as beta-carbonic anhydrases and a bicarbonate transporter, were significantly altered by both elevated CO2 and growth rate, which is considered to be a response to down-regulation of CCM genes (Hennon et al. 2019). This suggests that raphidophytes may have the genetic capacity to use bicarbonate through its own CCM.
ACKNOWLEDGEMENTS
We would like to thank Professor Hae Jin Jeong for kindly offering Chattonella ovata strain and the Culture Collection of KIOST for providing Chattonella marina strain. This research was supported by a National Research Foundation (NRF) grant funded by the Korean government (MSIT) (NRF-2016R1A6A1A03012647, NRF-2020R1A2C3005053) to K.Y.K.
SUPPLEMENTARY MATERIALS
Supplementary Fig. S1. Continuous monitoring of temperatures for each temperature-controlled aquarium up to 10 d under ambient (A & B) and elevated pCO2 (D & E) conditions for Chattonella marina and C. ovata, and an additional experiment for the growth rate of C. marina and C. ovata at the optimal growth temperature of 30°C under ambient and elevated pCO2 (C & F) condition (https://e-algae.org).
Supplementary Fig. S2. Growth curves of Chattonella marina over time. Each point represents mean ± standard deviation (n = 3) for ambient (●) or elevated pCO2 (▲) treatments in combination with 13, 20, 26, 30, or 34°C (https://e-algae.org).
Supplementary Fig. S3. Growth curves of Chattonella ovata over time. Each point represents mean ± standard deviation (n = 3) for ambient (●) or elevated pCO2 (▲) treatments in combination with 13, 20, 26, 30, or 34°C (https://e-algae.org).
Supplementary Fig. S4. Maximum efficiency of photosystem II (PSII) photochemistry (Fv/Fm) of cultures at ambient (A & B) and elevated pCO2 (C & D) levels for Chattonella marina and C. ovata at five temperatures (13, 20, 26, 30, 34°C) during the experiment. Error bars represent mean ± standard deviation (n = 3) (https://e-algae.org).