Statistical optimization of phytol and polyunsaturated fatty acid production in the Antarctic microalga Micractinium variabile KSF0031
Article information
Abstract
Polar microorganisms produce physiologically active substances to adapt to harsh environments, and these substances can be used as biomedical compounds. The green microalga Micractinium variabile KSF0031, which was isolated from Antarctica, produced phytol, a natural antimicrobial agent. Furthermore, several polyunsaturated fatty acids (PUFAs), including omega-3, exhibit antioxidant properties. Here statistical methods (Plackett-Burman design and Box-Behnken design) were used to optimize the culture medium of KSF0031 to improve biomass production, and K2HPO4, MgSO4·7H2O, and ammonium ferric citrate green (AFCg) were selected as significant components of the culture medium. Changes in the concentration of K2HPO4 and MgSO4·7H2O as positive factors and AFCg as a negative factor affected cell growth to a remarkable degree. The biomass production in a 100 L culture using the optimized medium for 24 d at 18°C was improved by 37.5% compared to that obtained using the original BG-11 medium. The quantities of PUFAs and phytol obtained were 13 mg g−1 dry cell weight (DCW) and 10.98 mg g−1 DCW, which represent improved yields of 11.70% and 48.78%, respectively. The results of this study could contribute to an improved production of phytol and fatty acids from Antarctic microalgae in the biomedical industry.
INTRODUCTION
The Antarctic and Arctic are polar regions with a year-round cold climate. Antarctica exhibits some of the coldest temperatures on earth because the supply of warm heat is interrupted by strong winds and the Antarctic circumpolar current (Barker et al. 2007). Psychrotrophic and psychrophilic microorganisms, which are adapted to cold environments, produce various physiologically active substances, such as cold-active enzymes and ice-binding proteins (Borchert et al. 2017, Brouchkov et al. 2017, Kim et al. 2017, Park et al. 2018). Polar microalgae also have mechanisms to survive in low-temperature environments. In a recent study, a novel ice-binding protein derived from polar microalgae was reported (Cho et al. 2019). Moreover, to survive at low temperatures, polar microalgae accumulate polyunsaturated fatty acids (PUFAs) inside their cells, which increase membrane fluidity; the flexible cell membrane reduces physical damage caused by the growth of ice crystals (Teoh et al. 2004). Unsaturated fatty acids have lower melting points than saturated fatty acids. This is due to their chemical structure, which includes multiple double bonds (Siram et al. 2019). PUFAs are valuable compounds, which include linoleic acid, gamma-linolenic acid, eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA). EPA and DHA, representative omega-3 unsaturated fatty acids, are essential for maintaining health (Gutiérrez et al. 2019). The PUFAs of polar microalgae, can be used as food additives and nutrients (Kim et al. 2016, Remize et al. 2021). In addition, ethanol extracts from the Antarctic freshwater microalgae Chloromonas reticulata KSF0100 and Micractinium sp. KSF0105 have been reported to exhibit anticancer and anti-inflammatory activities (Suh et al. 2018, 2019).
Phytol is a member of the diterpene alcohol family and a precursor of vitamins E and K1. Phytol is a component of chlorophyll, which is produced by all photosynthetic organisms, such as green algae, other plants, and some bacteria. Phytol has been reported to show anticancer activity, immunity enhancement, and antimicrobial effect (Komiya et al. 1999, Prabuseenivasan et al. 2006, Chowdhury et al. 2013, Murbach Teles Andrade et al. 2014). Phytol exhibits antibacterial and antifungal activities, high stability, and low toxicity; therefore, it is useful as a surface disinfectant (Ghaneian et al. 2015). Chemical synthesis is the primary method of industrial-scale phytol production; however, it is expensive and causes environmental pollution (Islam et al. 2015). The production of biopharmaceuticals, such as phytol, using microalgae could be an environment-friendly and sustainable method that could lower the production costs while reducing greenhouse gases, as it utilizes sunlight and atmospheric CO2 (Khan et al. 2018). In this study, we report, for the first time, that Micractinium variabile KSF0031, a species recently discovered in the Antarctic (Chae et al. 2019), accumulates significant quantities of phytol and PUFAs. Here, we optimized its medium to improve the production of cell biomass and bioactive ingredients.
MATERIALS AND METHODS
Strain and culture media
The Antarctic freshwater microalga Micractinium variabile KSF0031 was isolated from Ardley Island, South Shetland Islands, Antarctica (Chae et al. 2019). The culture was maintained in BG-11 medium (Stanier et al. 1971). The BG-11 medium contained the following (mM): NaNO3, 17.60; K2HPO4, 0.23; MgSO4·7H2O, 0.30; CaCl2·2H2O, 0.24; citric acid, 0.031; ammonium ferric citrate green (AFCg), 0.021; EDTA·Na2, 0.0027; Na2CO3, 0.19; trace metals, 0.058 (H3BO3, 0.046; MnCl2·4H2O, 0.009; ZnSO4·7H2O, 0.077; Na2MoO4·2H2O, 0.0016; CuSO4·5H2O, 0.0003; Co(NO3)2·6H2O, 0.00017).
Culture conditions
Ten liters of seed with approximately 5 × 105 cells mL−1 was inoculated in 100 L of BG-11 medium at 18°C (Supplementary Fig. S1) and cultured for 24 d. The KSF0031 was irradiated with cool white fluorescent light (80 μmol photon m−2 s−1) on a 16 : 8 h light-dark cycle with filtered atmospheric air supplied at a rate of 5 L min−1.
Statistical optimization
The essential components for the growth of KSF0031 were analyzed using the Plackett-Burman design (Plackett and Burman 1946). A classic way to screen for large variables is to use the Plackett-Burman design, which is programmed to identify critical parameters using n + 1 experiments (Ekpenyong et al. 2017). Based on these results, five elements of the BG-11 medium were selected and represented at two levels, low (−) and high (+) concentrations, producing a first-order model. The effect of each factor on cell numbers was calculated using Eq. (1):
, where Y denotes the response (number of cells), β0 is the model intercept, βi is the linear factor coefficient, and Xi is the level of each variable. The concentrations of the medium components were optimized using the Box-Behnken surface response method (Box and Behnken 1960). The Box-Behnken design was devised for analyses where each factor varies among the three levels (−1, 0, +1) (Czyrski and Sznura 2019). Accordingly, the individual factors in this study were coded into three disparate levels: (−) for low, (0) for intermediate, and (+) for high concentrations. A second-order polynomial model was designed to investigate the influence of individual components on the predicted response. A second-order polynomial model was calculated using Eq. (2):
, where Y denotes the predicted response (number of cells), β0 is the constant coefficient, βi, βij, and βii denote the regression coefficients of the model, and Xi and Xj represent the coded values of independent nutrient variables.
Cell growth monitoring and assay of fatty acid methyl esters and phytol
The growth of KSF0031 was analyzed by counting the cells using a hemocytometer and an optical microscope (Zeiss Axio Imager A2, Gottingen, Germany). Twenty milligrams of freeze-dried KSF0031 samples was used to extract fatty acids and phytol. Fatty acid methyl esters (FAMEs) were converted from crude lipids and analyzed as described previously (Kim et al. 2016). FAMEs and phytol were quantified using an external standard (1 mg of C22:0 in hexane); qualitative analysis was performed using a gas chromatography–mass spectrometer (TSQ 8000 EVO GC-MS; Thermo Fisher Scientific, Waltham, MA, USA) and reconfirmed using authentic standards (Supelco 37 Component FAME Mix and phytol; Sigma-Aldrich, St. Louis, MO, USA). The content (w w−1, %) and yield (mg L−1) were summed and expressed in terms of milligrams of FAMEs (or phytol) per gram of dry cell weight (DCW) and milligrams of FAMEs (or phytol) per liter of culture volume.
Statistical tests
Results with a p-value < 0.05 were considered statistically significant. The optimization values of significant medium components were calculated using the Minitab software ver. 14 (Minitab Inc., State College, PA, USA) and linear multiple regression was used for the statistical analysis.
RESULTS AND DISCUSSION
Component selection
To eliminate unnecessary or negative elements of the microalgal medium, all nine components were analyzed individually (Fig. 1). In the elimination test to optimize the culture of KSF0031, we found five significant factors among the medium components, namely K2HPO4, MgSO4·7H2O, citric acid, AFCg, and Na2CO3, all of which had positive effects. In each medium, when any one of these five components were removed, the growth of KSF0031 was restricted compared to that observed upon using the original BG-11 medium. The non-significant factors were NaNO3, CaCl2·2H2O, EDTA·Na2, and trace metals. In each medium, when any of these four components were removed, the growth of KSF0031 was not significantly affected when compared to that observed upon using the original BG-11 medium (Fig. 1).
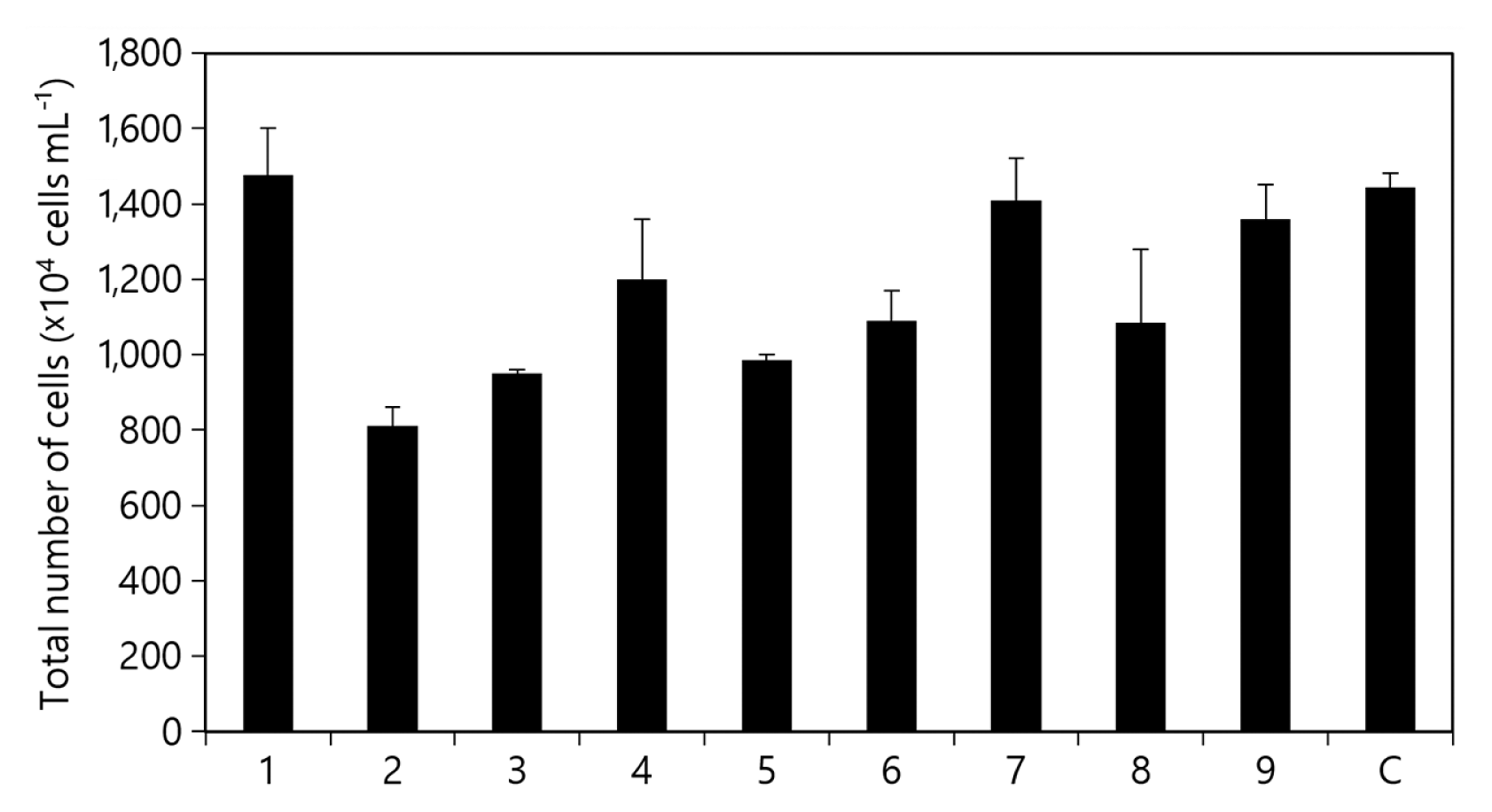
Elimination test results for KSF0031. Effect of each component on cell growth. Total cell numbers in the absence of 1, NaNO3; 2, K2HPO4; 3, MgSO4·7H2O; 4, CaCl2·2H2O; 5, citric acid; 6, ammonium ferric citrate green; 7, EDTA·Na2; 8, Na2CO3; 9, trace metals; C, control (no absence). Data shown represent the average cell densities ± standard deviation from triplicates (n = 3).
Screening of components using the Placket-Burman design
To select the component having optimal effects on KSF0031 cell numbers, the five components showing positive effects were tested using the Plackett-Burman design. Table 1 lists the test values, effects, t-statistics, and p-values. Results with a p-value lower than 0.05 were treated as showing a significant effect on the growth of KSF0031. NaNO3, CaCl2·2H2O, EDTA·Na2, and trace metals were not significant factors (p > 0.05) (Table 1). The remaining components (K2HPO4, MgSO4·7H2O, citric acid, AFCg, and Na2CO3) were significant (p < 0.05). These components were re-analyzed using a secondary Plackett-Burman design. Ten media, in each of which the concentrations were adjusted to -value (mM) and +value (mM), were prepared for the component analysis corresponding to the five significant variables (Table 2). Cell growth was measured in media adjusted to each concentration. Experiments were performed by varying the concentration range (0.2 times [−], 1.1 times [0], and 2 times [+] of the original concentration of BG-11) of the significant factors obtained through the elimination test, and the number of cells was measured. Three components showed significant effects, where two components, K2HPO4 and MgSO4·7H2O, exhibited positive effects; AFCg exhibited a negative effect (Table 2).

Statistical analysis of medium component using the initial Plackett-Burman test
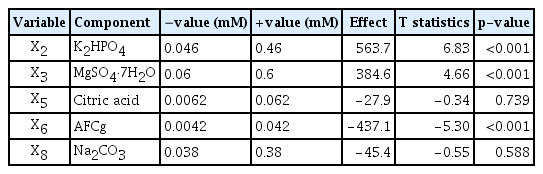
Statistical analysis of selected medium components using the secondary Plackett-Burman design
Optimization of medium components for KSF0031 growth
The Box-Behnken design was performed based on the statistical results of the Plackett-Burman design of KSF0031. It was applied to measure the interdependence of three component variables, K2HPO4 (X2), MgSO4·7H2O (X3), and AFCg (X6), in their effects on cell biomass. For positive effect factors, (+) was set as twice the original concentration (mM) of BG-11 medium, and (−) was set as 1/10 of (+). For negative effect factors, (+) was set as the original concentration (mM) and (−) was set as 1/10 of (+). Other factors (NaNO3, CaCl2·2H2O, citric acid, EDTA·Na2, Na2CO3, and trace metals) were added at their original concentrations (mM) (Table 3).
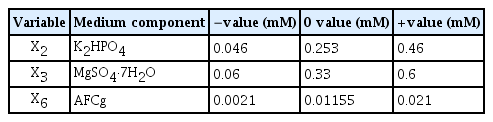
Box-Behnken optimization of selected significant medium components
The optimization values were calculated (Table 3). The estimated coefficients, t-statistics, and p-values were also calculated (Table 4). The effects of the other three variables at different levels are shown in Fig. 2. The expected number of KSF0031 cells was calculated using the Minitab software and determined to be 1.90 × 107 cells mL−1 with optimized concentrations of K2HPO4, MgSO4·7H2O, and AFCg as 0.4, 0.55, and 0.0021 mM, respectively. The final composition was as follows (mM): NaNO3, 17.6; K2HPO4, 0.4; MgSO4·7H2O, 0.55; CaCl2·2H2O, 0.24; citric acid, 0.031; AFCg, 0.0021; EDTA·Na2, 0.0027; Na2CO3, 0.19; and trace metals, 0.058 (Table 5).
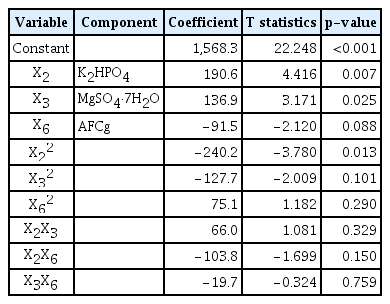
Response surface regression for selected medium components using Box-Behnken design
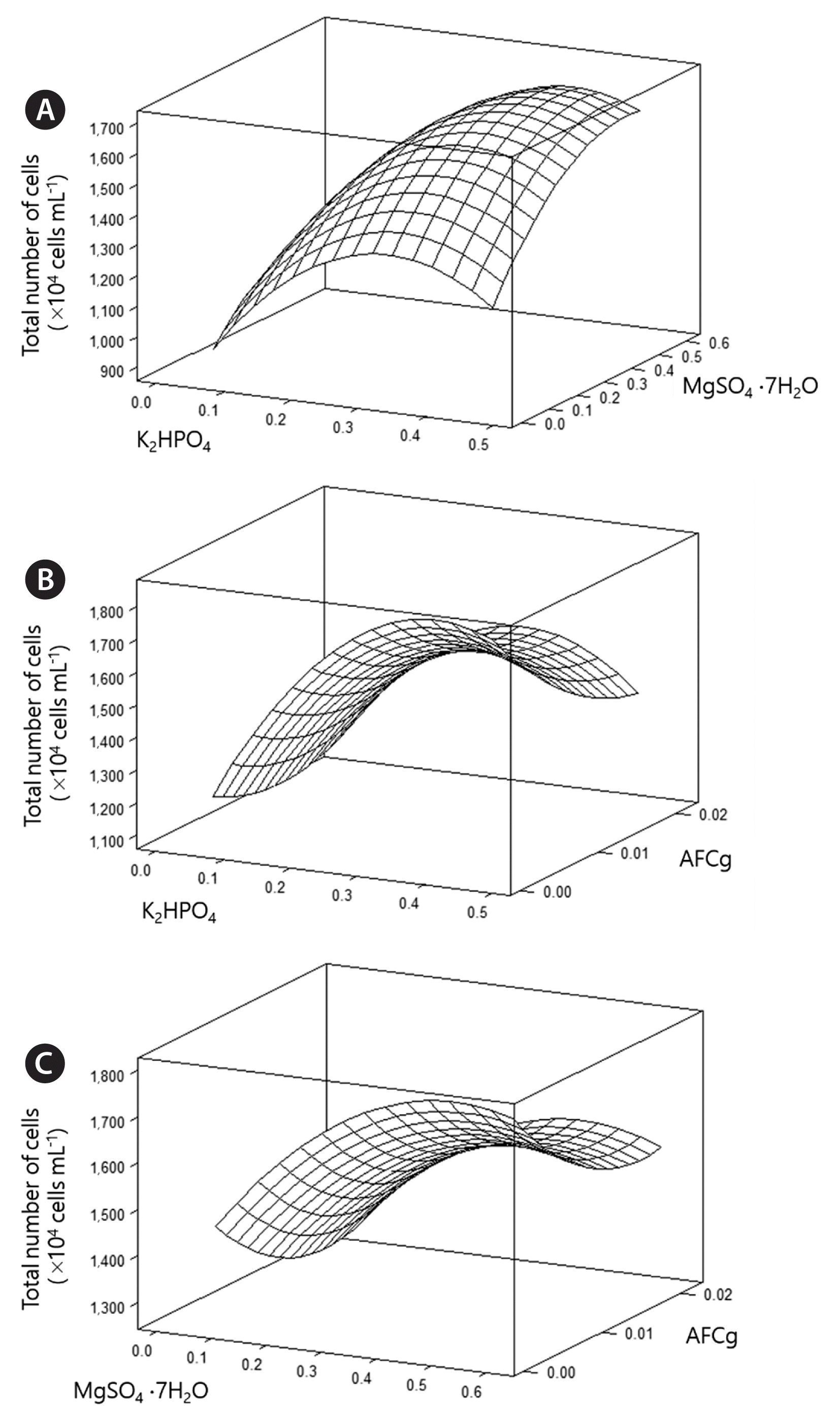
Three-dimensional response plots showing the effects of K2HPO4 and MgSO4·7H2O (A), K2HPO4 and ammonium ferric citrate green (AFCg) (B), and MgSO4·7H2O and AFCg (C).
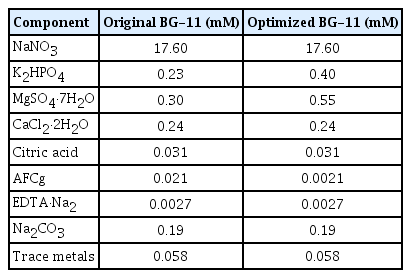
Comparison of medium composition of original BG-11 and optimized BG-11
Comparison of KSF0031 growth, and lipid and phytol production in the original medium vs. optimized medium
The growth of KSF0031 and its production of lipids and phytol in the medium optimized via the Box-Behnken design were analyzed. Fig. 3B shows the number of KSF0031 cells mL−1 in 100 L BG-11 culture media for 24 d. Maximal concentrations of (160 ± 8) × 105 and (225 ± 11) × 105 cells mL−1 were obtained in the original and optimized BG-11 medium, respectively. The dry mass of harvested cells cultivated in the optimized BG-11 medium was 35.60 ± 1 g, which constituted a 37.5% improvement in biomass when compared with the dry mass of KSF0031 cells from the original BG-11 medium (Fig. 3C). Although the levels of the two components were elevated in the optimized medium, the production cost of KSF0031 in this medium could be significantly reduced (Supplementary Table S1). The biomass of KSF0031 was increased in the optimized medium where the concentration of the negative factor AFCg was reduced by 10 fold. This gain was due to relief from iron toxicity effects on the cells. Iron, which can be considered a heavy metal, plays an important role in metabolism at low concentrations but can be toxic to microalgal cells at high concentrations (Subramaniyam et al. 2016). Through statistical optimization, it is possible to determine the appropriate concentration of heavy metal ions (Jaafari and Yaghmaeian 2019). After cultivation for 24 d in the original and optimized media, lipids were extracted from the dried biomass to obtain FAMEs and phytol. The PUFA content in the total fatty acids of Micractinium reisseri was 66% (Abou-Shanab et al. 2014). The PUFA content of Micractinium singularis reported in 2020 was 55.5% (Jo et al. 2020). There was no information on PUFA production (mg g−1 DCW) reported in either of these studies. In a study with M. reisseri reported in 2019, only 0.54 mg g−1 DCW of PUFA was produced (Elshobary et al. 2019). Reports on PUFA and phytol production in Micractinium species are limited, and research on the applications of Micractinium sp. is in its infancy. To evaluate the production of PUFAs and phytol, gas chromatography analysis was performed (Fig. 4, Supplementary Fig. S2). The dominant fatty acids of KSF0031 were linolenic acid (C18:3) and hexadecatrienoic acid (C16:3). Sorokina et al. (2020) reported that the dominant fatty acids in Micractinium sp. IC-44 were C16:0 and C18:2 when cultured in Bold’s basal medium at 28°C. Although the two strains belong to the same genus, KSF0031 is a psychrotrophic microalga in Antarctica, which has the characteristic of producing more PUFAs (Teoh et al. 2004). Our data indicate that compared to the original BG-11, using the optimized BG-11 medium leads to higher FAME and phytol production values (13.03 and 10.98 mg g−1 DCW, representing 12.04 and 48.78% yield increases, respectively) (Table 6). Phytol-producing microalgal strains have rarely been reported; the phytol contents of the ethanol extracts of Porphyridium cruentum, Phaeodactylum tricornutum, and Chlorella vulgaris were 3.1, 8.4, and 5.5%, respectively (Rodriguez-Garcia and Guil-Guerrero 2008). There have been no studies on phytol production using polar microalgae, and here, we performed an optimized medium design for the first time. The Antarctic microalga KSF0031 showed a high phytol content of 33.29% of total fatty acids, and we were able to improve the phytol content to 39.95% by using optimized BG-11 medium.

Comparison of cell growth and biomass of KSF0031 in different conditions in 100-L culture. (A) 100-L photobioreactor for 1 d (left) and 24 d (right). (B) Time profile of KSF0031 cell growth. The open and solid circles represent the original and optimized BG-11 conditions, respectively. (C) Cell weights with moisture removed. Data are represented as mean ± standard deviation of duplicates (n = 2).
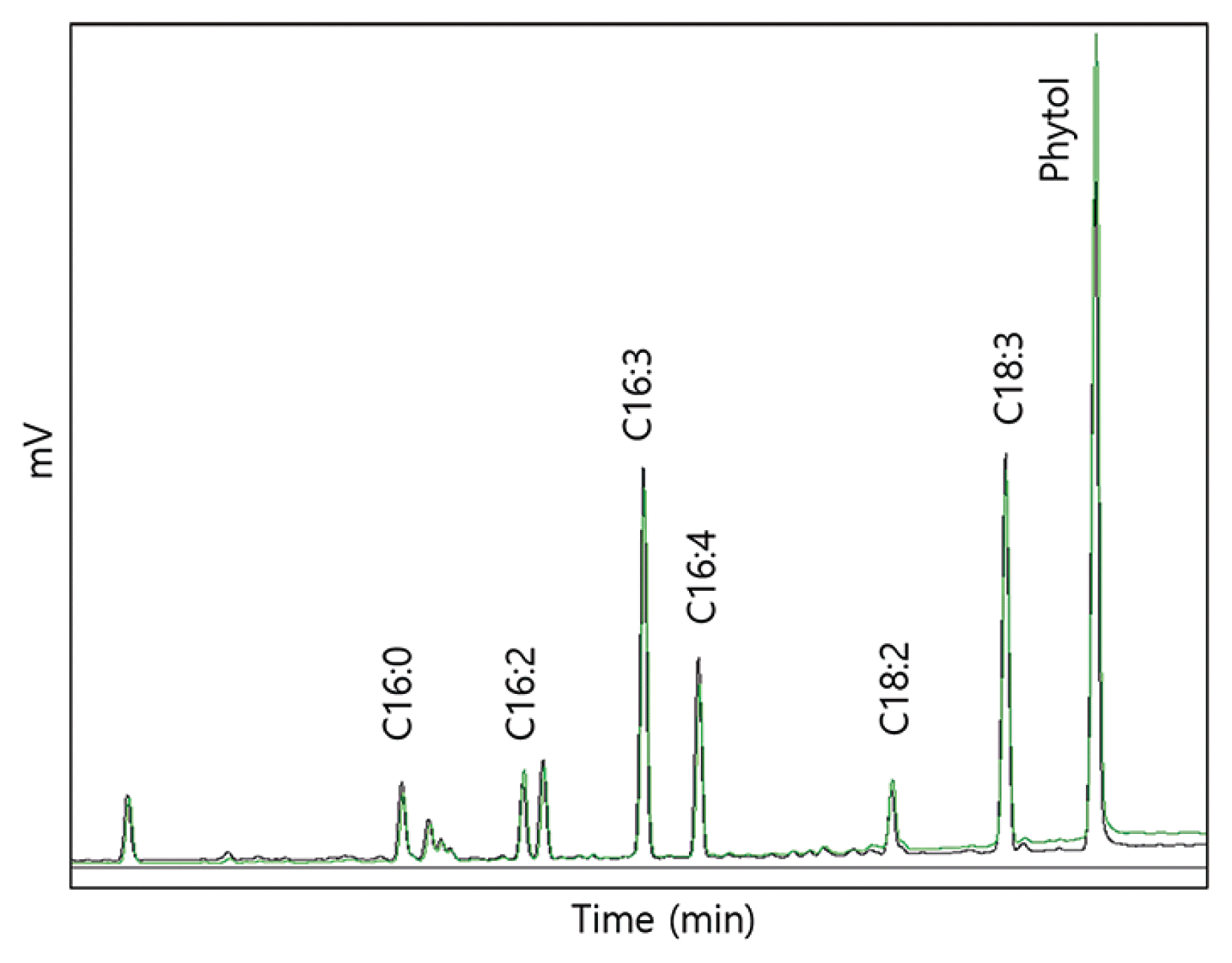
Gas chromatography results showing the fatty acid methyl esters and phytol on extracts of Micractinium variabile KSF0031. Chromatograph peak: black line, extract in original BG-11; green line, extract in optimized BG-11 on KSF0031.
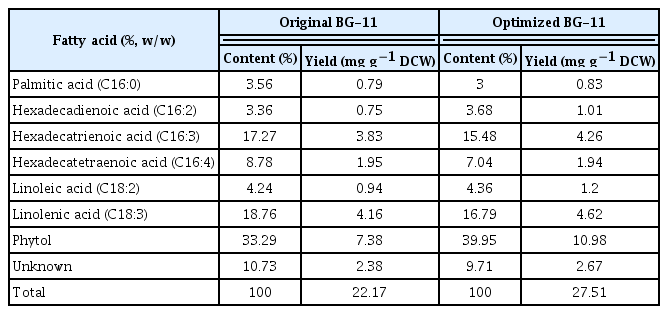
Content and yield of FAMEs and phytol from KSF0031
CONCLUSION
Here, we reported the production of phytol and PUFAs as bioactive compounds in the Antarctic microalga Micractinium variabile KSF0031 for the first time; further, we optimized their production. To enhance the biomass productivity of KSF0031, we statistically optimized the medium components using the Plackett-Burman and Box-Behnken designs. When AFCg, a component that had a negative effect, was diminished and K2HPO4 and MgSO4·7H2O, components that had a positive effect, were elevated, the biomass production of KSF0031 was increased. In this study, a 37.5% increase in biomass production was obtained using a sequential statistical method. The use of the optimized medium was shown to reduce the production cost of KSF0031. In addition, we obtained 12.04% and 48.78% improved PUFA and phytol levels, respectively, in the optimized BG-11 medium. Because this Antarctic microalga could be a potential resource in the biomedical industry, the medium designed in this study may play an important role in improving the production of cell biomass and bioactive ingredients.
SUPPLEMENTARY MATERIALS
Supplementary Table S1. Comparison of costs of media used for the biomass production of KSF0031 (https://www.e-algae.org).
Supplementary Fig. S1. Effect of temperature on the growth of Micractinium variabile KSF0031 (https://www.e-algae.org).
Supplementary Fig. S2. Gas chromatography mass spectrometry results showing the fatty acid methyl esters and phytol with extracts of Micractinium variabile KSF0031 (https://www.e-algae.org).
ACKNOWLEDGEMENTS
The research was supported by Grants from the Korea Polar Research Institute (PE22140 and PE19910).
Abbreviations
AFCg
ammonium ferric citrate green
DHA
docosahexaenoic acid
EPA
eicosapentaenoic acid
FAMEs
fatty acid methyl esters
PUFAs
polyunsaturated fatty acids
Notes
The authors declare that they have no potential conflicts of interest.