Tropical red alga Compsopogon caeruleus: an indicator of thermally polluted waters of Europe in the context of temperature and oxygen requirements
Article information
Abstract
The red alga Compsopogon caeruleus can generally be found in tropical and subtropical waters worldwide. In addition to its natural habitats, this species may be found in waters that receive abnormally hot water, e.g., from powerhouses. To date, the presence of C. caeruleus has not been observed in thermally polluted lacustrine ecosystems in Poland, which has a moderate climate. The thalli of this red alga were found growing on Vallisneria spiralis in Licheńskie Lake. Importantly, this paper presents a previously unknown relationship between the temperature (20, 25, 30, 35, and 40°C) and oxygen requirements of C. caeruleus (based on ex situ measurements of O2 consumption by thalli). Surprisingly, 35°C can be the optimum temperature for C. caeruleus, and this temperature is higher than the values reported by some previous thermal analyses by approximately 10°C. Additionally, we reviewed and mapped the distribution of this nonnative and mesophilic red alga in natural / seminatural water ecosystems in Europe. Finally, we propose that the occurrence of C. caeruleus mature thalli can be a novel, simple and easy-to-recognize bioindicator of artificially and permanently heated waters in moderate climate zones by a regular discharge of postindustrial water.
INTRODUCTION
The red alga Compsopogon caeruleus (Balbis ex C. Agardh) Montagne 1846: 154 (as ‘coeruleus’) is a widespread species in freshwater as well as marine tropical and subtropical ecosystems (Guiry and Guiry 2022). This macroalga has often been described as a native species in the Americas, e.g., USA (Florida) and Brazil, Asia, e.g., India, Africa, e.g., Algeria, and the Pacific Islands, e.g., USA (Hawaiian Islands) (Starmach 1977, Guiry and Guiry 2022). In addition, C. caeruleus populations have been found in ecosystems outside their natural range, such as in rivers and channels in Central Europe, which contain thermally contaminated waters due to discharge from factories and powerhouses (Fig. 1, Supplementary Table S1) (Weiss and Murray 1909, Täuscher 2012, Žáková et al. 2013). However, this nonnative and, according to us, potentially invasive species is best ecologically suited to the southern part of the European continent. Based on a report published a decade ago, C. caeruleus is present in only a few independent stands in Europe (Eloranta and Kwandrans 2012). In our opinion, this perspective requires substantial extension and revision.
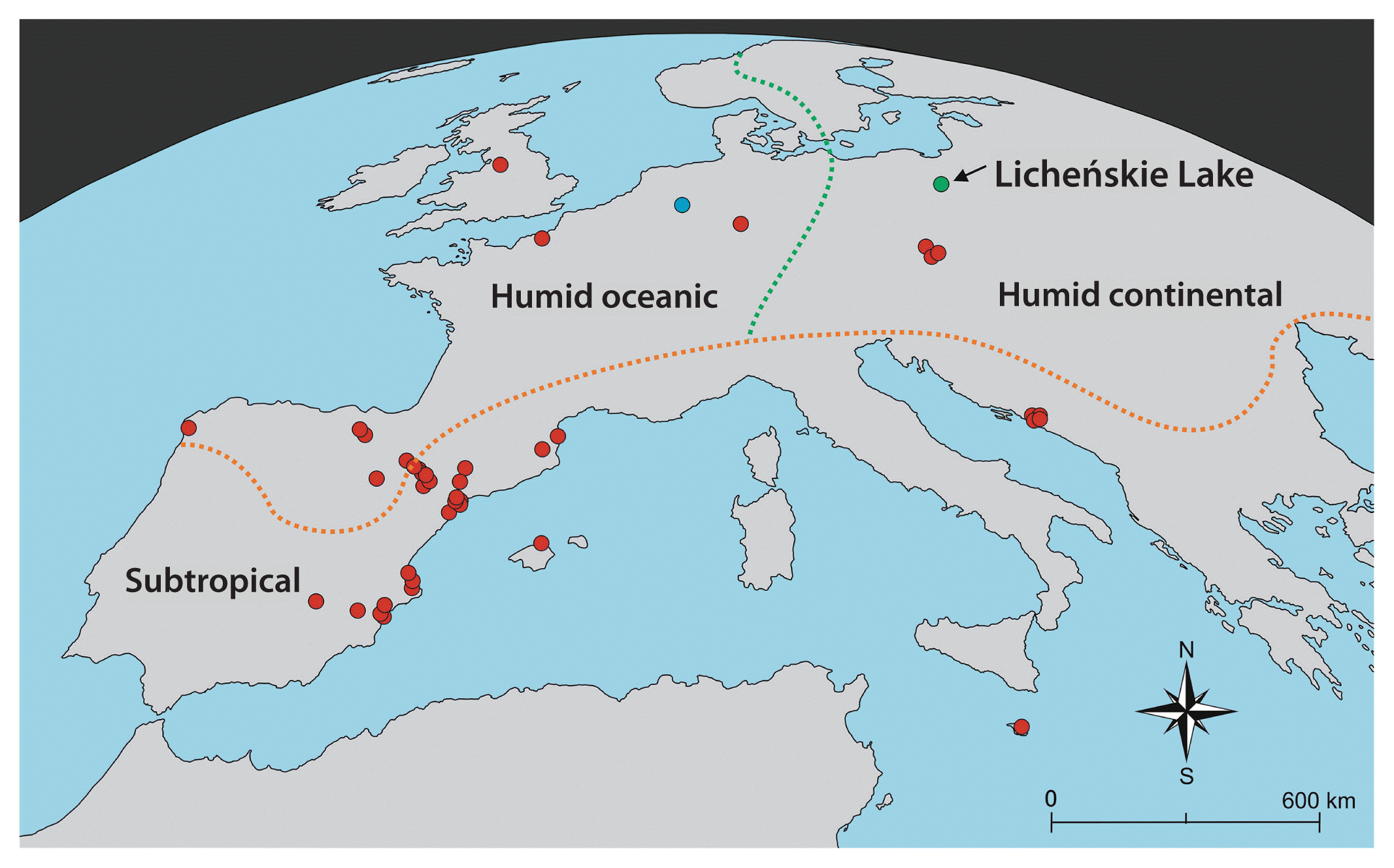
Distribution of Compsopogon caeruleus in Europe. Red circles, stands in natural and seminatural habitats reported in the literature and herbaria materials; green circle, the new stand from Licheńskie Lake (Poland); blue circle, location in the Eire River (Germany) where the reference material was collected. The dotted lines (orange and green) indicate the range of climate zones (Köppen 1900, Beck et al. 2018).
Moreover, artificial systems in Europe, e.g., garden ponds, glass / palm houses, and public / private aquaria with tropical animals and vascular plants, can host C. caeruleus (Starmach 1978, Battiato et al. 1979, Szymańska and Krupińska 1983, Gärtner 1987, Stoyneva et al. 2006). Although these reports incidentally called these systems “new stands”, we disagree with this classification, as these locations seen as artificial “enclaves” are entirely irrelevant for biogeographical studies of C. caeruleus, including studies on the expansion, population dynamics and distribution patterns of this species. However, from such an artificial system, nonnative tropical species, including C. caeruleus, may “escape” into the local environment (Zhan et al. 2021). Importantly, such a scenario is only possible if this kind of species encounters favorable conditions in a given country, allowing gradual acclimatization and further spreading. To date, in Central Europe, stable populations of C. caeruleus, which have survived the winter, have been recorded only in constantly thermally contaminated water ecosystems (Weiss and Murray 1909, Täuscher 2012, Žáková et al. 2013, 2015).
The structure of the C. caeruleus thallus is very characteristic among red alga and usually shaped like a horse’s tail, but the color of the thallus varies from dark olive, greenish, and bluish to violet–green (Krishnamurthy 1962, Starmach 1977, Eloranta and Kwandrans 2007, 2012, Sheath and Vis 2015). C. caeruleus is most often a benthic alga (attaches to water plants, macroalgae or stones with rhizoids or basal discs), but sometimes, it can be found in free-floating forms (Starmach 1977, Zhan et al. 2021).
The genus Compsopogon Montagne 1846 belongs to the family Compsopogonaceae (order: Compsopogonales, class: Compsopogonophyceae, subphylum: Proteorhodophytina, phylum: Rhodophyta) (Guiry and Guiry 2022). Historically, the Compsopogon genus was thought to include approximately 20 species (Guiry and Guiry 2022); however, as a result of the molecular revision, it was found that most of the organisms belong to the same species, namely, C. caeruleus (Balbis ex C. Agardh) Montagne 1846 (Necchi et al. 2013, Nan et al. 2016). Therefore, the following synonyms of C. caeruleus are used: C. hookeri Montagne 1846, C. chalybeus Kützing 1849, C. aeruginosus (J. Agardh) Kützing 1849, C. leptoclados Montagne 1850, C. lividus De Toni 1897, C. oishii Okamura 1915, C. minutus C. C. Jao 1941, C. iyengarii Krishnamurthy 1958, C. indicus Das 1963, C. lusitanicum M. P. Reis 1977, C. corticrassus Chihara & Nakamura 1980, C. aegyptiacus A. A. Aleem 1981, C. prolificus Yadava & Kumano 1985, C. tenellus Y. J. Ling & S. L. Xie 1998, C. aeruginosus var. catenatum Yadava & Pandey 1980, Compsopogonopsis leptoclados (Montagne) V. K. Krishnamurthy 1962 and Compsopogonopsis japonica Chihara 1976. Currently, within the genus Compsopogon, there are only six different species with taxonomically accepted names. Among these six species, C. argentinensis Pujals 1967, C. corinaldii (Meneghini) Kützing 1857, C. lividus De Toni 1897, C. occidentalis Tracanna 1980, and C. sparsus S. L. Xie & Y. J. Ling 1998 were recorded only in freshwater habitats, while C. helwanii El-Gamal & Salah El-Din 1999 was associated with sulphate springs (Guiry and Guiry 2022). C. caeruleus, the most common species in both hemispheres, has the widest range of tolerance to salinity and thus is found in freshwater, brackish and marine habitats (Guiry and Guiry 2022). In natural and seminatural ecosystems, populations of C. caeruleus develop mainly in flowing waters with temperatures above 20°C, high oxygenation and conductivity, and neutral pH (Tomás et al. 1980, Eloranta and Kwandrans 2007, Koletić et al. 2020b).
This work aims to update the understanding of the distribution of C. caeruleus in the water ecosystems of Europe, taking into account new findings and reviewed ecological data covering over 100 years. Moreover, in a novel assessment, we performed an ex situ characterization of how a temperature gradient (20–40°C) influences oxygen consumption by the thalli of C. caeruleus.
Our results fundamentally change the current view regarding the optimal temperature requirements and biogeography of this red alga, and this report describes a new and first stand of C. caeruleus in Poland (currently the northernmost and easternmost area of Europe where this species has been found). The potential application of C. caeruleus as an indicator of thermally polluted waters is highlighted.
MATERIALS AND METHODS
Morphological identification and nomenclature
The morphological identification of red alga samples relied on examining the features of the thalli according to taxonomical keys (Kwandrans and Eloranta 2010, Eloranta et al. 2011, Sheath and Vis 2015). Our specimens from Licheńskie Lake were initially identified as C. caeruleus (Balbis ex C. Agardh) Montagne 1846: 154 (as ‘caerulaeus’). The nomenclature of the Compsopogon taxa presented here has been verified to comply with AlgaeBase (Guiry and Guiry 2022).
Molecular identification
For molecular identification, genomic DNA was isolated from Compsopogon thalli collected in Licheńskie Lake, both fresh samples and prepared herbarium specimens. To confirm our morphological identification of C. caeruleus, we used sequence data of the mitochondrial cytochrome c oxidase subunit I (COI) and the nuclear region of SSU rRNA (V9) gene fragments. The COI gene fragment (328 bp) was amplified using bcdF01 (CATTTTCHACTAAYCATAARGATATTGG) and bcdR06 (GGDGGRTAHACAGTYCAHCCNGT) primers (Trzebny et al. 2020). The V9 (130 bp) fragment was amplified using 1391F (GTACACACCGCCCGTC) (Lane 1991) and EukBr (TGATCCTTCTGCAGGTTCACCTAC) (Medlin et al. 1988) primers. The polymerase chain reaction primers were fused with dual-indexed Ion Torrent adapters. Amplicon sequencing was performed using Ion S5 sequencing system (Thermo Fisher Scientific, Waltham, MA, USA) on 530 Ion chip according to the manufacturer’s protocols. Sequence reads were analyzed using a custom workflow in Geneious Prime v2022.2.2 (Biomatters Ltd., Auckland, New Zealand). Sequences generated in this study and concerning C. caeruleus from Licheńskie Lake were published in GenBank under accession numbers OP727285 (COI) and OP737800 (V9). Both fresh and deposited in herbarium material of examined red alga from Licheńskie Lake shared the same COI and V9 sequences. Furthermore, the sequence of COI generated in our study was identical to those obtained from C. caeruleus collected in Argentina (GenBank, accession No. MH638996), China (KY083068), and the USA, Hawaii (KC596325). In turn, V9 sequences revealed 100% identity with C. caeruleus recorded in Saint Lucia (AF087127) and in the USA, Hawaii (KC596284) and Florida (AF087124).
Biogeography of Compsopogon caeruleus in Europe
The distribution of C. caeruleus in Europe was analyzed using different data sources, such as databases, i.e., DAISIE, GBIF, and WRiMS (Roy et al. 2022, The Global Biodiversity Information Facility 2022, World Register of Introduced Marine Species 2022), records from herbaria (Beaty Biodiversity Museum, Algae Collection 2022, National Museum of Natural History, Botany Collection 2022, The New York Botanical Garden, C. V. Starr Virtual Herbarium 2022), and published papers (see Supplementary Table S1). Climate zones are defined according to the updated Köppen-Geiger classification (Köppen 1900, Beck et al. 2018).
Lacustrine location of Compsopogon caeruleus
The samples of the studied red alga were collected from a stand in the artificially heated waters of Licheńskie Lake (52°18′56.9″ N, 18°21′05.0″ E; Licheń, Greater Poland) in June 2021 and September 2022. Licheńskie Lake is a seminatural ecosystem and belongs to the Konin-Pątnów powerhouse cooling system (Table 1, Fig. 2). This waterbody is eutrophic (according to Carlson indices), monomictic with a volume of 6,712.3 m3, has a water surface of approximately 147 ha, and has a mean depth of 4.5 m (Table 1) (Pyka et al. 2007, Ejsmont-Karabin and Hutorowicz 2011, Bogacka-Kapusta and Kapusta 2013, Ejsmont-Karabin et al. 2020, Zdanowski et al. 2020). In this thermally polluted lake (where the average water temperature of the epilimnion in summertime ranges from 25 to 28°C) (Table 2), Vallisneria spiralis L. (commonly known as tape grass) is the dominant macrophyte and forms dense phytocoenoses along the shoreline (Gąbka 2002, Hutorowicz 2006, Ejsmont-Karabin and Hutorowicz 2011, Bogacka-Kapusta and Kapusta 2013, Ejsmont-Karabin et al. 2020). The thermal pollution of Licheńskie Lake is defined as deviation from the natural temperature in a habitat resulting from elevated temperature associated with industrial cooling activities (Dodds and Whiles 2010).
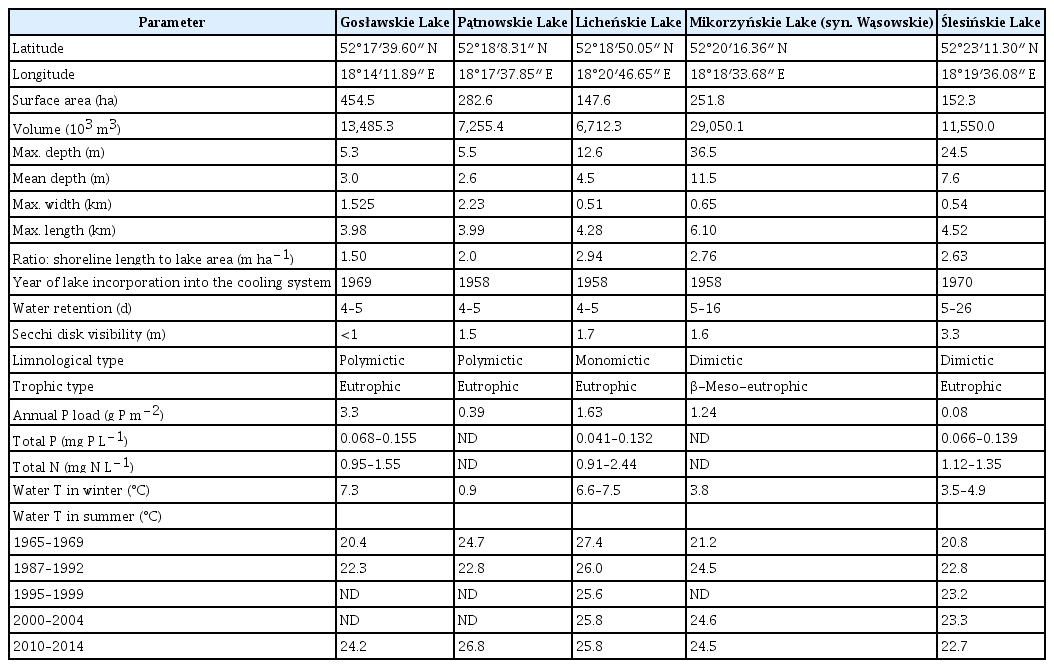
Morphometric and thermal data of the Konin Lakes included in the cooling water system of coal-fired Pątnów-Konin powerhouses
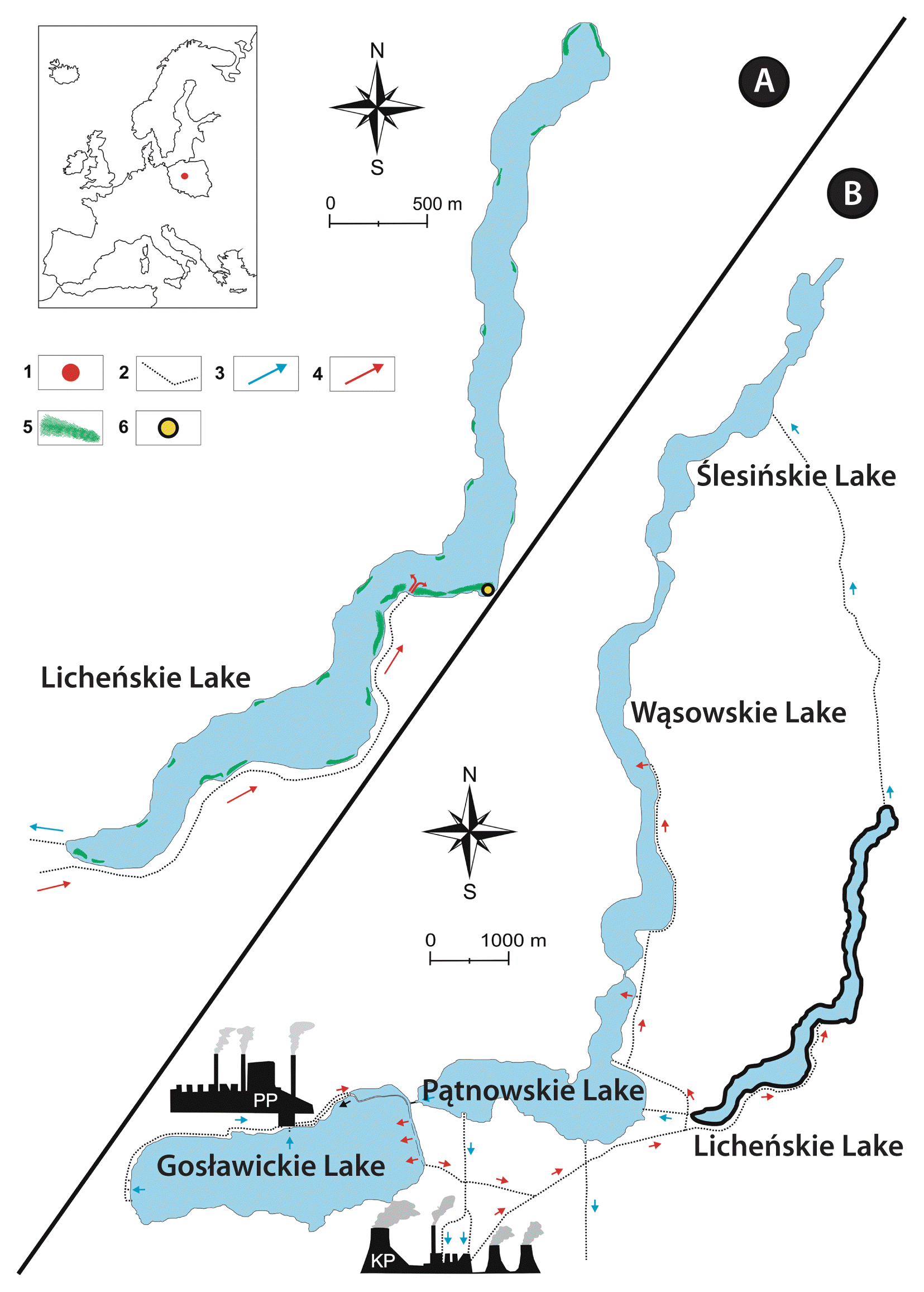
(A) Licheńskie Lake in detail. (B) Schematic of the water cooling system for two powerhouses at Konin Lakes. Blue arrows, intake of cooling waters; red arrows, discharge of heated waters; PP, Pątnów powerhouse; KP, Konin powerhouse; 1, location of the new Compsopogon caeruleus population in Europe; 2, channels of the cooling system; 3, cold water flow direction; 4, heated water flow direction; 5, Vallisneria spiralis meadows; 6, stand of C. caeruleus in the lake.

Summer epilimnion water temperatures (mean values, with the range in parentheses) of Licheńskie Lake from 1965–2018 (Ejsmont-Karabin et al. 2020)
Culture of Compsopogon caeruleus
The thalli of C. caeruleus were solely found on the leaves of tape grass in Licheńskie Lake and then stored under laboratory conditions. Wright’s cryptophyte (WC) medium (pH 7.3, adjusted with NaOH; CaCl2·2H2O, MgSO4·7H2O, NaHCO3, K2HPO4, NaNO3, Na2SiO3·9H2O, Na2·EDTA, FeCl3·6H2O, CuSO4·5H2O, ZnSO4·7H2O, CoCl2·6H2O, MnCl2·4H2O, Na2MoO4·2H2O, H3BO3), vitamins (thiamin HCl, and biotin), and n-tris(hydroxymethyl)methyl-2-aminoethanesulfonate (TES buffer) were used to culture the algae (Guillard and Lorenzen 1972) in a Conviron CMP 6050 phyrotron (Winnipeg, MB, Canada) (25°C, 12 L : 12 D, 250 lux, and 50% humidity).
Herbarium
Several individuals of C. caeruleus were selected as herbarium samples. A herbarium sheet (voucher code: POZ-A-COM2021-01) was prepared following the standard protocol (Drobnik 2007) and digitalized. The thallus samples were attached to a labeled sheet using Archer’s method. A voucher specimen (in a Merrill case) was deposited in the Natural History Collections at the Faculty of Biology (Poznań Algae Herbarium [POZA]; 52°28′00.8″ N, 16°55′29.3″ E) at Adam Mickiewicz University, Poznań, Poland.
Water samples from Compsopogon caeruleus habitat
Water samples were taken directly from the Vallisneria underwater meadow using 1.0 L sterile plastic bottles (Roth, Karlsruhe, Germany). Long-sleeved veterinary-grade gloves were used during sampling to prevent contamination. The water samples were filtered through a plastic sieve to eliminate small vascular plants (e.g., lemnids), filamentous green algae (e.g., Rhizoclonium, Cladophora and Ulva) and other objects (e.g., fragments of decaying wood). Subsequently, the water samples were placed in two (0.5 L) sterile plastic containers (Roth), and the first subsample was preserved using 0.5 mL of chloroform (Sigma-Aldrich, Seelze, Germany) and cooled at 4°C. Later, this preserved sample was purified in the laboratory through a microbiological filter with a pore size of 0.45 μm and stored in a freezer at −20°C for further chemical analyses. The second subsample of water (0.5 L), which was used to measure pigment concentration (chlorophyll a), was not prefiltered with a microbiological filter.
Physicochemical properties of water
In the field (at the C. caeruleus site), water temperature, pH, electrolytic conductivity, total dissolved solids (TDS), and oxygenation were measured using the Professional Plus multiparameter instrument (YSI, Yellow Springs, OH, USA). A Secchi disk was used to assess water transparency. Water depth was measured using a plastic staff gauge.
The chemical analyses, including analyses of ammonium (NH4+), nitrite (NO2−), nitrate (NO3−), phosphate (PO43−), sulphate (SO42−), and total phosphorus (P), nitrogen (N), and iron (Fe ions) concentrations, as well as water color analyses, were performed at the laboratory using a HACH DR 2800 spectrophotometer (Fairborn, OH, USA) according to standard hydrochemical methods (Baird et al. 2017). The concentrations of calcium (Ca2+), magnesium (Mg2+), potassium (K+), sodium (Na+), and chloride (Cl−) were determined by high-performance liquid chromatography (Shimadzu, Kyoto, Japan). The sodium chloride (NaCl) and calcium carbonate (CaCO3) concentrations were measured with the use of the HACH Chloride and Calcium Carbonate Digital Titrator Kit. The turbidity of the water was measured by a turbidimeter EUTECH TN-100 (Thermo Scientific, Singapore). The chlorophyll a concentration (corrected for pheopigments) was determined fluorometrically (Beutler et al. 2002).
Morphological analysis
The morphology of the C. caeruleus thalli was assessed as soon as possible after samples were collected from Licheńskie Lake. Macroscopic observations were performed using a stereomicroscope (Stemi DV4; Carl Zeiss, Göttingen, Germany). The cells were analyzed using a light microscope (Axioskop 2 MOT; Carl Zeiss). Photographs of fresh samples and fragments of thalli (that had been stained for 15 min with Lugol’s solution) were captured with CapturePro 2.77 software (Jenoptic, Jena, Germany).
Reference material
Comparative material for examining oxygen requirements under different temperatures and molecular analysis was obtained from the Culture Collection of Algae (SAG) at Göttingen University (Compsopogon hookeri Montagne 1846: 157, strain number SAG 37.94). The thalli of Compsopogon that had originally been collected in 1994 from the Erft River (Bedburg, Germany; 50°59′18.0″ N, 6°34′36.5″ E) were cultured in MiEB12 and SWES media (in a 10 : 1 proportion). Importantly, the Erft River carries heated wastewater from a sugar refinery (SAG 2022). According to the taxonomic review (including molecular methods) of the genus Compsopogon, the species C. hookeri is now considered a synonym of C. caeruleus (Balbis ex C. Agardh) Montagne 1846: 154 (Guiry and Guiry 2022).
Temperature gradient and oxygen consumption
Oxygen uptake was measured polarographically with a Clark-type oxygen electrode (Hansatech Instruments, King’s Lynn, UK) at different temperatures, i.e., 20, 25, 30, 35, and 40°C, in 0.7 mL of the incubation medium. It must be highlighted that before the oxygraph experiments, C. caeruleus thalli were kept at 25°C, which was determined to be the optimum temperature for this species in nature for some European stands (Koletić et al. 2020b). The temperature stability in the oxygraph chamber was maintained by a water jacket from LAUDA ECO heating thermostats (Lauda-Königshofen, Germany) with a bath (to maintain an accurate temperature of the fresh respiration buffer). For C. caeruleus from Licheńskie Lake, WC medium (see above) was used as the incubation medium. In turn, C. caeruleus, which was purchased from the SAG collection, was examined in a mixture of MiEB12 and SWES media (in a 10 : 1 proportion). The purine nucleotides ATP, ADP, GTP, and GDP were purchased from Sigma-Aldrich. The respiratory rate at steady state (VO) is expressed in nmol O min−1 g−1 of thallus. The oxygen concentrations in the incubation media were comparable and stable across the temperature gradient used (approximately 7.3 mg O2 L−1) and measured with a ProDSS Multiparameter Digital Water Quality Meter, which was equipped with a ProDSS optical dissolved oxygen sensor (YSI). The light intensity during these experiments was measured by a digital luxmeter LX1010BS (Yanmis, China) and reached 210 lux.
Statistical analysis
The oxygraph results are presented as the means ± standard deviation from five independent experiments, with each measurement performed at least in duplicate. Statistically significant differences between the measurements across the temperature gradient were established with unpaired two-tailed Student’s t-test, and significance levels are indicated as follows: p < 0.05 (*), p < 0.01 (**), or p < 0.001 (***).
Literature acquisition
The literature search was performed using databases such as Elsevier Science Direct (2022), Google Scholar (2022), JSTOR (2022), Mendeley Data (2022), PubMed (2022), RCIN (2022), Scopus (2022), and SpiringerLink (2022).
RESULTS
Stand, morphology, and ecology of Compsopogon from Poland
The presence of C. caeruleus has not been officially recorded in seminatural or natural habitats in Poland (outside closed artificial habitats such as aquaria). The new stand of C. caeruleus in Central Europe was located along the shoreline of Licheńskie Lake. The thalli of this red alga grew on the long leaves of the tropical vascular plant V. spiralis in summertime (Fig. 3A). The morphology of lacustrine C. caeruleus is consistent with the outline of previously described specimens from other freshwater habitats (see above). The thalli were greenish blue and strongly branched, especially in the middle part, with a length of 1 mm to 15 cm and a width of filaments 50–2,000 μm. The uniseriate central axis contained flat cells with a length of 50–140 μm and a width of 50–250 μm (Fig. 3G). The central axis of the thallus was constricted at the base and attached to the substratum with the help of the conical basal holdfast (Fig. 3B). The cortex consisted of polygonal or irregular cells (15–40 μm × 20–40 μm) (Fig. 3H) and formed colorless rhizoids in the middle and basal parts of filaments (Fig. 3C & D). The cortical cells contained several discoid-shaped, parietal, and olive-green chloroplasts (Fig. 3H). The young uniseriate filaments of lateral branches (without cortex layers) consisted of discoidal or cylindrical cells that were apically rounded (Fig. 3F). Monosporangia with an irregular shape were present mainly in the upper parts of the thalli and were 15–22 μm in diameter (Fig. 3H).

Habitat and thalli of Compsopogon caeruleus from Licheńskie Lake. (A) Meadow of tape grass (white circle, thalli of C. caeruleus on Vallisneria spiralis leaves). (B) Thalli with conical basal holdfast (black arrow). (C) Basal part of thalli with visible colorless rhizoids (black arrows). (D) Cortex of thallus with (on the left side) and without (on the right side) rhizoids. (E) Irregular cells of the cortex. (F) Young uniseriate filament without cortex. (G) Central axis consisting of flattened cells. (H) Cortex layer with monosporangium (black arrow). (I) Details of the cortex layer. Scale bars represent: B & C, 200 μm; D & E, 100 μm; F, 50 μm; G–I, 20 μm.
The C. caeruleus thalli we studied were found in water with a mean temperature of 27.6°C, basic pH (8.6), and proper aerobic conditions (average oxygenation level: 79%, average oxygen concentration: 6.4 mg L−1) (Table 3). The water was greenish (with a chlorophyll a concentration of 21.5 μg L−1), but the sandy bottom was completely visible to a depth of 50 cm. The water color was 25 Pt-Co units on average, and the turbidity ranged from 18 to 23 nephelometric turbidity units (NTU). Notably, high concentrations of CaCO3 (283.2 mg L−1) and SO42− (77.1 mg L−1) were salient features of the C. caeruleus habitat, which had average concentrations of mineral compounds. Therefore, moderate levels of conductivity (617.3 μS cm−1) and TDS (375.2 mg L−1) were observed. The chemical analysis showed that the concentrations of nutrients were high (PO43−, 0.1 mg L−1; NH4+, 0.2 mg L−1; and NO3−, 0.9 mg L−1) (Table 3).

Physicochemical and biological properties of the water (mean values, with the range in parentheses) from the stand of Compsopogon caeruleus in Licheńskie Lake (n = 10)
Habitats of Compsopogon caeruleus in Europe
Based on the available literature published in several languages over the 100 years that we analyzed, C. caeruleus populations in Europe were generally found in natural and seminatural ecosystems with flowing waters such as rivers (60% of cases) and channels (18%). Less commonly, this red alga was found in springs (5%), coastal lagoons (5%), lakes (4%), ponds (4%), and streams (4%). As many as 73% of all known C. caeruleus habitats were categorized as freshwater, 13% as brackish water, 9% as freshwater / brackish water, and 5% were at the transition between brackish and saline waters (Supplementary Table S1).
C. caeruleus belongs to the group of organisms known as phytobenthos, and our analysis indicates that this red alga is mainly an epiphytic species; it has been recorded on the leaves, shoots and roots of many vascular aquatic plants, including Arundo donax, Ceratophyllum demersum, Phragmites australis, Potamogeton crispus, P. pectinatus, Ranunculus fluitans, Sparganium emersum, and V. spiralis (Supplementary Table S1). Moreover, C. caeruleus may also grow on the thalli of other macroalgae, mainly those from the phylum Chlorophyta (i.e., Cladophora sp., Pithophora sp., and Rhizoclonium sp.), and Charophyta (Chara sp.) (Supplementary Table S1). Additionally, C. caeruleus is an epilithic alga and can attach to natural hard clay and sandy bottoms or rocks, stones, pebbles, and artificial materials (concrete walls of rivers or streams that are submerged in water). Moreover, solid organic substrates, such as shells of water snails and underwater wood (dead trees and shrubs), constitute microhabitats of C. caeruleus (Supplementary Table S1).
We noticed that from the ecological and phytosociological perspective, the thalli of C. caeruleus are elements of the communities of submerged, nonrooted floating, rooted floating, and emergent plants. That is, C. caeruleus was found in macroalgal associations, such as Charetum asperae, Charetum balticae, and Charetum hispidae (Supplementary Table S1). Moreover, its thalli can be found in water habitats dominated by pondweed species (i.e., Potamogeton pectinatus, P. pusillus, P. nodosus, and P. coloratus). Other associations, i.e., Parvopotamo-Zannichellietum, Myriophylletum spicati, M. verticillati, Ceratophylletum demersi, Lemnetum trisulcae, and Nupharo-Nymphaeetum albae, are also environmental niches for C. caeruleus (Supplementary Table S1).
Ecological requirements
Considering all the stands of C. caeruleus mentioned above, the highest temperature of water in which this red alga was found was 32.2°C, with a median of 22.9°C (Supplementary Table S1). The most important physicochemical parameters of its habitats are as follows: (1) water depth, 20–100 cm; (2) water flow, 0.3–48 m3 s−1; (3) pH, 6.8–8.7; (4) conductivity, 48.6–38,200 μS cm−1; (5) TDS, 375.5–3,990 mg L−1; (6) water color, 24–12,789 Pt-Co mg L−1; (7) salinity, 0.4–4.6 PSU; (8) turbidity, 1.1–23 NTU; (9) alkalinity, 42.5–295 mg CaCO3 L−1; (10) dissolved oxygen, 2.4–13.1 mg L−1; (11) NO3−, 0.15–23 mg L−1; (12) NH4+, 0.03–3.2 mg L−1; (13) PO43−, 0.0047–8.7 mg L−1; and (14) SO42−, 43.2–3,341.8 mg L−1 (Supplementary Table S1).
Temperature affects oxygen consumption by Compsopogon caeruleus thalli
Among the five different temperatures tested, i.e., 20, 25, 30, 35, and 40°C, the maximal respiratory rate of C. caeruleus thalli at a steady state (VOmax) was achieved at two temperatures, namely, 35 and 40°C, with rates of 228.24 ± 20.44 and 231.34 ± 22 nmol O min−1 g−1 of thallus, respectively (Fig. 4). The lowest VO was observed at 20°C (48.12 ± 6.58), but the rate was just over double the value at 25°C (108.1 ± 9.7). Interestingly, the dark respiration rate can also be approximately doubled for this species after a shift from 20 to 25°C (Necchi and Zucchi 2001). At 30°C, another increase in the VO was observed (128.4 ± 10.66), and this increase almost doubled after the transition to higher temperatures (35 and 40°C). This trend was also observed for the reference C. caeruleus from SAG with very similar values of VO and VOmax. Our results fit the general observation for plants where the respiratory rate can be doubled per 10°C rise in temperature (Q10 = 2) (Atkin and Tjoelker 2003). However, we also observed that a 5°C rise in temperature can lead to this type of change in VO. This means that the acceleration of oxygen metabolism may depend on starting temperature, i.e., for C. caeruleus, the shift from 20 to 25 or 30 to 35°C resulted in an approximate doubling in VO, but the other doubling in VO occurred for the change from 25 to 35°C. Again, it fits the Q10 interpretation in the context of VO changes, as the value of this factor is not constant and influenced by the temperature range (Atkin and Tjoelker 2003). Targeting nucleotide metabolism combined with oxygen metabolism, we used exogenous nucleotides such as ATP, ADP, GTP, and GDP (up to 2 mM). However, the addition of these nucleotides did not influence the VO regardless of the temperature.
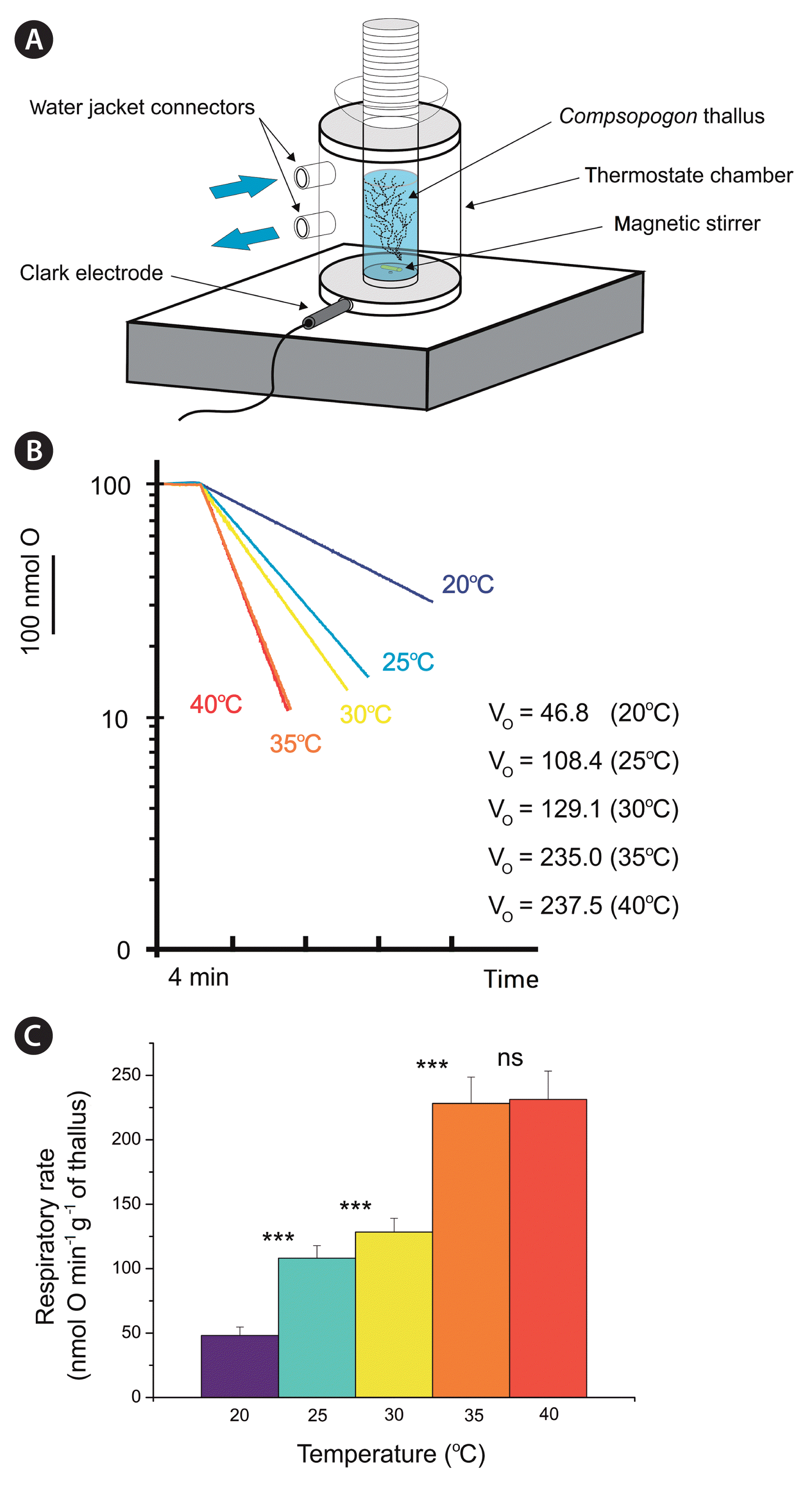
(A) Schematic representation of the oxygraph chamber with a water jacket and Clark-type electrode. (B) Representative oxygraphic traces (series of technical repetition) of Compsopogon caeruleus thalli (from Licheńskie Lake) exposed to different temperatures (oxygen content on the x-axis is exponential, where 100 indicates full air saturation and 0 indicates anoxia). (C) The effect of temperature change on respiratory rate at steady state (VO) expressed in nmol O min−1 g−1 of C. caeruleus thallus. ns, non-statistically significant; ***, statistically significant (p < 0.001).
DISCUSSION
Currently, the presence of C. caeruleus, a nonindigenous European algal flora species (with presumable origins in South America), has been reported mainly in the southern and southwestern parts of the continent (Fig. 1, Supplementary Table S1) (Koletić et al. 2020b, Guiry and Guiry 2022). In particular, the natural and seminatural freshwater and brackish riverine ecosystems of continental Spain are inhabited by C. caeruleus (Donze 1968, Tomás et al. 1980, 2013, Tomás 1981, 1988, Álvarez Cobelas 1984, Busquets et al. 1985, Aboal 1986, Aboal Sanjurjo 1989, Sabater et al. 1989, Aboal et al. 1994, Cantoral Uiza and Aboal Sanjurjo 2001, Edigos and Aboal 2003, Sánchez Castillo et al. 2011, Chapuis et al. 2014, García-Fernández et al. 2015). Stands of this alga in the Spanish Canary Islands have still not been reported, although the presence of this species in that area is mistakenly indexed in AlgaeBase (Guiry and Guiry 2022). In the area of the Mediterranean Sea, island populations of C. caeruleus were found in Mallorca (Balearic Islands, Spain) (Sánchez Castillo et al. 2011) and Malta (Maltese Archipelago, Europe) (Battiato et al. 1979). The Mediterranean basin’s subtropical climate favors this species in a few locations in France (originally identified as C. hookeri or C. aeruginosus [J. Agardh] Kützing) (Boillot 1958, Eloranta et al. 2011, Breton 2014) and Croatia (Koletić et al. 2019, 2020a, 2020b).
The earliest, i.e., from December 1908, record of C. caeruleus (originally identified as C. leptocladus) in a European location with a humid oceanic climate, namely, the United Kingdom (Manchester), concerns the Reddish Canal water (approximately 23°C), which was contaminated with artificially heated water from cotton factories (Weiss and Murray 1909); the species was observed again decades later (Fig. 1, Supplementary Table S1) (Blackler and Krishnamurthy 1961, Krishnamurthy 1962). The occurrence of all known populations of C. caeruleus found in Germany (some identified as C. hookeri Montagne) was also correlated with the postindustrial discharge of heated water (Friedrich 1966, 1973, Mauch et al. 2003, Hussner and Lösch 2005, Täuscher 2012). The same is true for Central European areas with a humid continental climate; in Austria and the Czech Republic, populations of C. caeruleus (first identified as C. aeruginosus [J. Agardh] Kützing) were observed in flowing-water habitats that were affected by thermal pollution from a chemical factory and nuclear powerhouse (Eloranta et al. 2011, Žáková et al. 2013). As the Reddish Canal in Manchester was closed many years ago, the currently described stand of C. caeruleus in Poland constitutes a novel record and the northernmost habitat of this species in Europe; the southernmost population of this red alga was found on Malta Island (Battiato et al. 1979).
Over the last 60 years, the biodiversity of Polish freshwater ecosystems has been profoundly influenced by the introduction of nonindigenous species (Najberek and Solarz 2011). A major biogeographical change occurred with the opening of coal-fired powerhouses, which typically use natural water (from nearby ponds, lakes, and rivers) for cooling installations. The discharge of heated water directly into the environment creates places that are called “invasion gates” for alien plant and animal species (Wilk-Woźniak and Najberek 2013). The Konin Lakes have been affected by this practice since 1958; thus, these lakes have been the object of eco-bioinvasive research and modeling for global warming changes (Zdanowski et al. 2020, Dziuba et al. 2021). This semiartificial lake system with heated waters (sometimes called “heated islands”) imitates the thermal conditions of tropical ecosystems; therefore, it can be easily colonized by alien organisms preferring warmer waters (Wilk-Woźniak and Najberek 2013). The Konin Lakes are inhabited by approximately 100 alien species, including vascular plants such as V. spiralis and Hygrophila polysperma (Indian swampweed) and animals such as Dreissena polymorpha (zebra mussel), Sinanodonta woodiana (Chinese pond mussel), and Pseudorasbora parva (topmouth gudgeon) (Gąbka and Owsianny 2009, Najberek and Solarz 2011, Zdanowski et al. 2020). The toxic bloom-forming cyanobacteria Raphidiopsis raciborskii (syn. Cylindrospermopsis raciborskii) and green microalgae Hariotina polychorda (syn. Coelastrum polychordum) have been reported among the microorganisms found in these lakes (Berger et al. 2006, Wilk-Woźniak and Najberek 2013). Alien species that achieve substantial biomass, e.g., tape grass and clams, and therefore modulate the structure and function of the Konin Lakes should be considered invasive. Unfortunately, people may treat Konin Lakes as a place where they can throw away unwanted tropical animals and plants. As a curiosity, some aquarium animals but alien species for Europe, such as herbivorous piranha (unpublished data without species details) and turtle, i.e., Trachemys scripta elegans known as the red-eared slider (personal observation), were noted in Konin Lakes. We can only presume how C. caeruleus was introduced into Licheńskie Lake, intentionally or accidentally. It is a very high probability that this red alga presence in Poland is a consequence of human activity.
No reports concerning the invasive potential of C. caeruleus, which inhabits artificially heated aquatic ecosystems of Europe, have been published to date. Currently, this red alga does not achieve substantial biomass in Licheńskie Lake. However, C. caeruleus, an epiphytic or epilithic species, is a competitor for native phytobenthos species such as micro- and macroalgae or animals, e.g., Rotifers, because it occupies habitat niches and exploits environmental resources such as nutrients, light, and oxygen. In this context, C. caeruleus has already modified or disrupted the ecosystem in which it began colonizing. Therefore, at this moment, we propose to use the term “potentially invasive species” for this red alga. The aspects and details of C. caeruleus synecology in semiartificial habitats of Europe require further research.
Notably, the bioindication potential of C. caeruleus is proposed here. Generally, species or communities that help to evaluate the quality of the environment affected by natural and / or anthropogenic changes over time are named bioindicators (Holt and Miller 2010, Parmar et al. 2016). For example, the presence of a given species in a stressed ecosystem is often correlated with the transmission of physicochemical factors such as temperature. According to the classification of bioindicators accepted by the World Conservation Union or International Union for Conservation of Nature in the context of our analyses, C. caeruleus may fit the subtype termed the “pollution bioindicator”. Our point of view is based on the fact that outdoor populations of C. caeruleus occur yearly and only in constantly thermally contaminated ecosystems of humid oceanic and humid continental climate zones of Europe (Fig. 1), i.e., C. caeruleus thalli are able to regenerate after possibly cold winters (December–March) but without a drop in water temperature below 0°C (Supplementary Table S1). More than 100 years ago, it was suggested that this red alga is prone to be winter-killed in an outdoor garden without heating based on observations in Great Britain (Colley 1916). The exact low and lethal water temperature for C. caeruleus has not been reported to date. In the Konin Lakes system, Ślesińskie Lake is the coldest (the average annual temperature of the water surface varies from 13.4 to 13.9°C), while Licheńskie Lake, in which C. caeruleus was found, is the warmest (the average annual temperature of the water surface varies from 15.6 to 16.6°C) (Tables 1 & 2, Fig. 1) (Ejsmont-Karabin et al. 2020). It must be noted that in Licheńskie Lake, even during the winter, the water temperature is never lower than 6.6°C (when powerhouses are working), and during the summertime, the surface water reaches temperature approximately 30°C (Table 2) (Zdanowski and Prusik 1994, Socha and Hutorowicz 2009, Najberek and Solarz 2011, Ejsmont-Karabin et al. 2020). Similarly, C. caeruleus was identified in the thermally polluted Dyje River in the Czech Republic during the vegetation period, which may reach only 0.8°C in winter (Žáková et al. 2013, 2015). Therefore, we can only surmise that the abovementioned low-temperature values make it possible for this species to survive in a given ecosystem in a resting form / stage. Accordingly, this red alga provides the bioindication potential for constant thermal pollution derived from human activity.
Moreover, our interpretation of C. caeruleus presence in some regions of Europe is fundamental for assessing the expansion of tropical algae outside the natural range. Importantly, the occurrence of C. caeruleus and its use as the bioindicator constitutes a considerable advantage compared to the manual / automatic measurement of water temperature from time to time. For example, during winter, the water temperature in Licheńskie Lake, which hosts C. caeruleus, may be as low as 6.6°C (Table 1). Such a temperature value measured directly by thermometer during warm winters in lakes / rivers of Poland, and other countries with a moderate climate, does not necessarily indicate permanent thermal pollution of the water ecosystem. In such cases, whether the measurement of water temperature is the result of weather conditions or temporary / constant thermal contamination may be ambiguous.
For many organisms, including algae, oxygen respiration is essential for maintaining metabolic homeostasis, and adequate nucleotide metabolism ensures survival (Woyda-Ploszczyca and Rybak 2021). Nucleoside diphosphate kinase (NDPK) is an important enzyme that regulates the pools of various nucleotides in cells. This transphosphorylase affects cell division by providing building blocks for the synthesis of DNA and RNA; for instance, this enzyme produces GTP via the following reaction: ATP + GDP → ADP + GTP. During the oxygraph measurement experiments, regardless of the temperature, C. caeruleus thalli did not respond to exogenously administered purine nucleotides (ATP, ADP, GTP, and GDP) as substrates or products related to NDPK. This could mean that the endogenous nucleotides were at saturating concentrations, and the cultivation medium fulfilled the requirements for proper growth. NDPK is a thermostable and well-conserved enzyme among different species (prokaryotic and eukaryotic), and the temperature of half denaturation for this kinase may reach 76°C; thus, 37°C can be the optimal temperature for the function of this kinase, while 60°C can completely inhibit its function (Miranda et al. 2008, Georgescauld et al. 2013). Therefore, our temperature gradient probably did not inhibit NDPK or other enzymes that are involved in nucleotide metabolism. It must be highlighted that unsuitable low or high temperatures (nonoptimal) may limit plants, including algae, development, e.g., growth rate, by decreasing enzyme activities / capacities and ultimately depleting ATP level below requirements (Raven and Geider 1988, Atkin and Tjoelker 2003, Nejrup et al. 2013, Ahmad et al. 2020).
The record presented in this paper substantially extends our knowledge about the eastward and northward distribution of C. caeruleus in Europe. The maximal water temperature of the seminatural habitats in which C. caeruleus has been found thus far is 32.2°C (Supplementary Table S1) (Žáková et al. 2013, 2015). However, the optimal temperature that supports the highest frequency of C. caeruleus, e.g., from natural ecosystems in Croatia (Neretva River valley), is approximately 25°C (Eloranta and Kwandrans 2007, Koletić et al. 2020b). A temperature of 25°C seems to be the most favorable for populations of C. caeruleus in non-European (Brazilian) regions, considering photosynthetic performance (Necchi 2006). Surprisingly, according to our ex situ polarographic measurements with the Clark-type oxygen electrode, the previously modeled thermal optimum for this red alga could be an underestimate. In our study, the shift from 35 to 40°C (independent measurements) did not result in further increases in the VO of C. caeruleus thalli, which can mean that 35°C is a borderline temperature for optimal development (Fig. 4). Therefore, the optimal temperature for growth of C. caeruleus may be increased by at least 10°C. Our results are strongly supported by laboratory findings that place the maximum photosynthesis rate of Compsopogon at 30–35°C (Kremer 1983, Necchi and Zucchi 2001, Sheath and Vis 2015). We are aware that regional ecotypes may differ in their environmental requirements, which explains variations in data concerning the optimum temperature for C. caeruleus. However, if 35°C is more optimal for the metabolic pathways of C. caeruleus, including respiration and photosynthesis, tested under laboratory conditions in different countries over nearly 40 years, namely, for populations of Poland (this paper) and Germany (Kremer 1983, Necchi and Zucchi 2001, Sheath and Vis 2015), this feature is relatively independent of geography. In this context, it could mean that even potential genetic variation between different populations did not affect the general metabolic strategy of this species. Such an interpretation is supported by our results concerning measurements of respiratory rates because regardless of C. caeruleus thalli origination, we observed very similar values of VO for a given temperature. Moreover, C. caeruleus from Licheńskie Lake revealed 100% identity with C. caeruleus from SAG, taking into account COI (OP747576) and V9 (OP737799) sequences.
Finally, we propose that C. caeruleus can populate warmer habitats, e.g., those affected by thermal pollution. Therefore, the presence of C. caeruleus in moderate climate zones can be a simple and easy-to-recognize (even without taking the thallus samples) indicator of artificially and permanently heated systems by a regular discharge of postindustrial water. Such conditions, combined with other environmental factors (e.g., nutrient levels), favor the expansion of alien species in a given region of Europe. The Konin Lakes can be a model for tracking the mechanisms of tropical / subtropical species adaptation into generally colder climatic zones. The growing demand for electricity and the resulting higher discharge of heated water may favor the development of this nonnative algae species in Europe. Interestingly, C. caeruleus can transiently survive even at 40°C (Fig. 4). Notably, only time will reveal how the distribution of this red alga will expand in response to an additional factor, i.e., an increase in the average annual temperature due to global warming. Importantly, changes in plant respiration influenced by temperature are currently seen as a crucial feature of the ecosystem response to global climate change (Atkin and Tjoelker 2003). Institutions responsible for environmental monitoring using phytobenthos as a bioindicator of the ecological state should include in their training programs and protocols information on the possibility of observing mesophilic macroalgae, such as C. caeruleus, in aquatic ecosystems.
ACKNOWLEDGEMENTS
We express our thanks to Urszula Sobczynska, Molecular Biology Techniques Laboratory, Faculty of Biology, Adam Mickiewicz University, Poznań, Poland, for her assistance in the molecular identification of the species. We are grateful to dr. Jarosław Kubiak from the Cartographic and Geodetic Environmental Research Laboratory, Faculty of Geographical and Geological Sciences, Adam Mickiewicz University, Poznań, Poland, for the recalculation of some Compsopogon geographical locations, i.e., the conversion from Universal Transverse Mercator (UTM) coordinates to a geographic coordinate system. This work was supported by the National Science Centre, Poland (grant 2015/19/D/NZ3/00087; principal investigator: dr. hab. Andrzej M. Woyda-Ploszczyca). The sponsor was not involved in preparing the article and deciding to submit the article for publication.
Abbreviations
ADP
adenosine diphosphate
ATP
adenosine triphosphate
COI
cytochrome c oxidase subunit I
GDP
guanosine diphosphate
GTP
guanosine triphosphate
NDPK
nucleoside diphosphate kinase
NTU
nephelometric turbidity units
TDS
total dissolved solids
VO
respiratory rate at steady state
VOmax
maximal respiratory rate at steady state
WC
Wright’s cryptophyte
Notes
The authors declare that they have no potential conflicts of interest.
SUPPLEMENTARY MATERIALS
Specification of Compsopogon caeruleus stands in natural and seminatural water ecosystems of Europe with ecological data (https://www.e-algae.org).