Estimation of bioluminescence intensity of the dinoflagellates Noctiluca scintillans, Polykrikos kofoidii, and Alexandrium mediterraneum populations in Korean waters using cell abundance and water temperature
Article information

Abstract
Many dinoflagellates produce bioluminescence. To estimate the intensity of bioluminescence produced by populations of the heterotrophic dinoflagellates Noctiluca scintillans and Polykrikos kofoidii and autotrophic dinoflagellate Alexandrium mediterraneum in Korean waters, we measured cellular bioluminescence intensity as a function of water temperature and calculated population bioluminescence intensity with cell abundances and water temperature. The mean 200-second-integrated bioluminescence intensity per cell (BLcell) of N. scintillans satiated with the chlorophyte Dunaliella salina decreased continuously with increasing water temperature from 5 to 25°C. However, the BLcell of P. kofoidii satiated with the mixotrophic dinoflagellate Alexandrium minutum continuously increased from 5 to 15°C but decreased at temperatures exceeding this (to 30°C). Similarly, the BLcell of A. mediterraneum continuously increased from 10 to 20°C but decreased between 20 and 30°C. The difference between highest and lowest BLcell of N. scintillans, P. kofoidii, and A. mediterraneum at the tested water temperatures was 3.5, 11.8, and 21.0 times, respectively, indicating that water temperature clearly affected BLcell. The highest estimated population bioluminescence intensity (BLpopul) of N. scintillans in Korean waters in 1998–2022 was 4.22 × 1013 relative light unit per liter (RLU L−1), which was 1,850 and 554,000 times greater than that of P. kofoidii and A. mediterraneum, respectively. This indicates that N. scintillans populations produced much brighter bioluminescence in Korean waters than the populations of P. kofoidii or A. mediterraneum.
INTRODUCTION
Many marine organisms produce visible light as bioluminescence (Widder 2010, Wilson and Hastings 2013). Dinoflagellates are ubiquitous and are one of the major components of marine ecosystems (Kang et al. 2023). Many dinoflagellate species produce bioluminescence (Haddock et al. 2010, Hinder et al. 2012, Cusick and Widder 2020, Jeong et al. 2021, Park et al. 2021). Dinoflagellates often dominate bioluminescence in surface waters, accounting for >90% of bioluminescence intensity measured in surface waters (Tett 1971, Lapota and Losee 1984, Lapota et al. 1988, Swift et al. 1995). The bioluminescence of dinoflagellates is used as a defense mechanism to deter predators and attract secondary predators (Burkenroad 1943, Esaias and Curl 1972, Haddock et al. 2010, Valiadi and Iglesias-Rodriguez 2013). The bioluminescence of dinoflagellates is usually produced by physical stressors, including waves, ships, and moving organisms (Esaias and Curl 1972, Morin 1983, Rohr et al. 1998, Latz et al. 2004, 2008, Maldonado and Latz 2007, Valiadi and Iglesias-Rodriguez 2013, Cusick and Widder 2014). Several bioluminescent dinoflagellates sometimes cause red tides or harmful algal blooms that produce spectacular luminous phenomena at night (Seliger et al. 1970, Wilson and Hastings 2013, Martínez et al. 2016, Detoni et al. 2023). Consequently, bioluminescence has attracted considerable attention from both military and ecological perspectives, playing a crucial role in submarine and ship detection, as well as the prediction of red tides and harmful algal blooms caused by bioluminescent plankton (Lynch 1981, Haddock et al. 2010, Le Tortorec et al. 2014, Cusick and Widder 2020).
Over the past 50 years, there have been several efforts on quantifying bioluminescence in the ocean (Lapota and Losee 1984, Lapota et al. 1988, Batchelder et al. 1992, Swift et al. 1995, Marcinko et al. 2013, Xue et al. 2020). One method to estimate the bioluminescence intensity of populations of a bioluminescent species is to multiply its abundance in a water parcel by the cellular bioluminescence intensity. The estimated bioluminescence intensities of populations of all bioluminescent species are often summed (Batchelder and Swift 1989, Buskey 1992, Swift et al. 1995). However, if any environmental variables affect the cellular bioluminescence intensity of a bioluminescent species, the cellular bioluminescence intensity should be measured as a function of those environmental variables.
Several environmental variables, including prey species, prey abundance, water temperature, and photoinhibition affect the cellular bioluminescence intensity of heterotrophic dinoflagellates (Jess 1985, Buskey et al. 1992, 1994, Buskey 1995, Latz and Jeong 1996, Li et al. 1996), while light intensity, nutrient concentrations, water temperature, salinity, and pH affect that of autotrophic dinoflagellates (Swift and Meunier 1976, Sweeney 1981, Jess 1985, Craig et al. 2003). For accurate estimation, investigations of the effects of additional environmental variables on the cellular bioluminescence intensity of dinoflagellates are required.
The heterotrophic dinoflagellate Noctiluca scintillans often causes the occurrence of red tides which glow at night (Padmakumar et al. 2010, Harrison et al. 2011, Lotliker et al. 2018, Valiadi et al. 2019, Temnykh et al. 2022). This dinoflagellate swims slowly and usually floats to the surface through large cell vacuoles filled with ammonium ions (Elbrächter and Qi 1998, Tiselius and Kiørboe 1998). Consequently, it forms very dense patches of red tides, seen as slicks, causing bright bioluminescence at night (Padmakumar et al. 2010, Temnykh et al. 2022). There has been one study on effects of water temperature on the cellular bioluminescence intensity of N. scintillans (Jess 1985). However, that study measured the cellular bioluminescence intensity of N. scintillans at only two water temperatures, 10 and 20°C but N. scintillans occurs across a wide range of water temperatures, approximately 2–30°C (Elbrächter and Qi 1998, Harrison et al. 2011, Ocean Biodiversity Information System 2024). Therefore, the cellular bioluminescence intensity of N. scintillans as a function of water temperature requires additional quantification.
The bioluminescent dinoflagellate Polykrikos kofoidii is ubiquitous (Buskey et al. 1992, Yoo et al. 2002, 2013, Tillmann 2004, Lim et al. 2017, Ocean Biodiversity Information System 2024) and an important predator of red-tide organisms (Matsuoka et al. 2000, Jeong et al. 2001, 2003, Kang et al. 2018, Kim et al. 2019). Only the duration of daylight has been explored as an environmental variable affecting the cellular bioluminescence intensity of P. kofoidii. Alexandrium mediterraneum has been found in the coastal waters of Greece, Italy, Spain, and Korea (John et al. 2003, Aguilera-Belmonte et al. 2011, Laza-Martínez 2017, Park et al. 2021), and recent research has reported its bioluminescent capability (Park et al. 2021). Any environmental variable affecting the cellular bioluminescence intensity of A. mediterraneum is yet to be investigated. Therefore, to better understand the bioluminescence of P. kofoidii and A. mediterraneum at seas, the effects of critical environmental variables affecting their cellular bioluminescence intensities should be explored.
In Korean waters, N. scintillans has been commonly found and sometimes causes red tides (National Institute of Fisheries Science, Korea 2024). P. kofoidii has also been commonly found in Korean waters and is often abundant after red tides (Yoo et al. 2013, Lim et al. 2017), while A. mediterraneum has been recently found (Park et al. 2021). The water temperature in Korean waters is 3–30°C (Real-time Information System for Aquaculture, National Institute of Fisheries Science 2024). Therefore, these three dinoflagellate species may experience a wide range of water temperatures, and their cellular bioluminescence intensity should be explored.
In the present study, the cellular bioluminescence intensity of N. scintillans, P. kofoidii, and A. mediterraneum as a function of water temperature was determined. From measurements of bioluminescence intensity and cell abundances of these three dinoflagellate species in Korean waters, population bioluminescence intensity was estimated. These results will provide a basis for understanding relationships between water temperature and the bioluminescence intensity of heterotrophic and autotrophic dinoflagellates and estimating their bioluminescence at seas.
MATERIALS AND METHODS
Preparation of experimental organisms
Cells of N. scintillans NSDJ2010 were isolated from coastal waters off Dangjin, Korea, and a clonal culture was established (Table 1). The cultures of N. scintillans were fed the chlorophyte Dunaliella salina as prey. For maintenance, dense cultures of N. scintillans (approximately 5–10 cells mL−1) were transferred to a 250-mL flask (Falcon; Corning Inc., New York, NY, USA) that contained prey species. Cultures of N. scintillans were incubated under 20 μmol photons m−2 s−1 of a light-emitting diode (LED; FS-075MU, 6500 K; Suram Inc., Suwon, Korea) and a 14 : 10 h light : dark cycle. Isolation, establishment, and maintenance (approximately 50–70 cells mL−1) of the culture of P. kofoidii PKJH1607 collected from coastal waters off Jangheung, Korea were similar to those for N. scintillans culture except that the mixotrophic dinoflagellate Alexandrium minutum was used as prey (Table 1). Cells of A. mediterraneum AMYS1807 were isolated from coastal waters off Yeosu, Korea, and a clonal culture was established (approximately 700–1,000 cells mL−1) (Table 1). The culture of A. mediterraneum was grown in F/2 seawater medium without silicate (Guillard and Ryther 1962) under 100 μmol photons m−2 s−1 of a LED and a 14 : 10 h light : dark cycle.
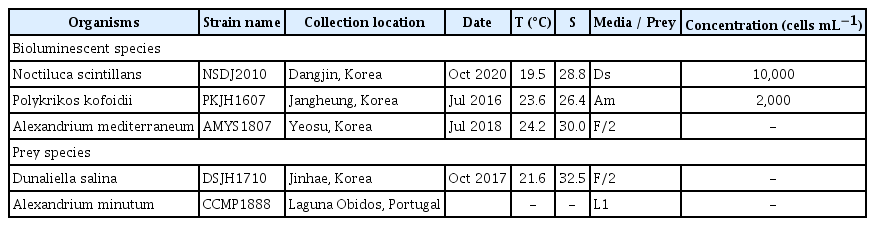
Information on strain name, isolation, and maintenance of the bioluminescent dinoflagellates and their prey species
Experimental setup
Experiments 1–3 were conducted to investigate the effects of water temperature (5 to 30°C) on the bioluminescence intensity of three bioluminescent species. The cultures were incubated constantly under the same condition or gradually transitioned to the target temperature across nine days to avoid any shock that may occur when a large change in the environment occurs too rapidly (Fig. 1). During the pre-incubation period, a 5-mL aliquot was subsampled from each flask at 2-day intervals and fixed with Lugol’s solution to determine the abundance of each species. All temperature experiments were conducted at salinity 30 under 20 μmol photons m−2 s−1 for N. scintillans and P. kofoidii and 100 μmol photons m−2 s−1 for A. mediterraneum at a 14 : 10 h light : dark cycle. If all cells in the flask were dead by increasing or decreasing temperature during the pre-incubation period, a further experiment at the water temperature was not conducted.

Water temperatures during pre-incubation and experimental incubation periods for each experimental organism (A–C). Dark blue colors indicate the period of incubation of cultures at the target water temperature. Bright yellow colors represent the period of temporarily maintaining the cultures at the water temperature two level lower or higher than target water temperature. Dark yellow colors represent the period of temporarily maintaining the cultures at the water temperature one level lower or higher than the target water temperature. This gradual acclimation minimizes any possible water temperature shock.
For Experiment 1, dense cultures of N. scintillans were transferred to six 250-mL flasks containing prey (D. salina) and filtered seawater. A dense prey culture was also transferred to six flasks containing F/2 medium. These cultures were acclimated gradually to the target temperature. However, all N. scintillans cells were dead at 30°C in the pre-incubation period; thus, the experiments were conducted only at 5, 10, 15, 20, and 25°C. After pre-incubation, three 1-mL aliquots were taken from each flask, and the abundance of each experimental species was determined using a light microscope (Olympus BX51; Olympus Co., Tokyo, Japan). When prey cells were undetectable in the acclimated N. scintillans culture, the initial concentrations of both predator and prey species were established by adding a predetermined volume of culture to each well of the experimental well plate (Table 2). The triplicate experimental wells with mixtures of both predator and prey species, triplicate prey control wells (D. salina only), and triplicate predator control wells (N. scintillans only) in 6-well culture plates were established.
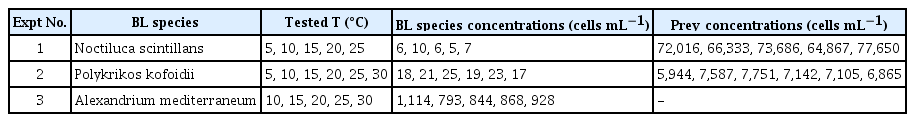
Experimental design
One milliliter of the F/2 medium adjusted to the target temperature was added to each well for each target temperature. A culture of N. scintillans with each target temperature was filtered with a 0.2-μm disposable syringe filter (DISMIC-25CS type, 25 mm; Advantec, Toyo Roshi Kaisha Ltd., Chiba, Japan). The filtrates were then added to the prey control well in the same volume as the predator culture added to the experimental well. Similarly, a culture of D. salina, adjusted to the target temperature, was filtered using a 0.2-μm disposable syringe filter, and the filtrates were added to the predator control well at the same volume as the prey culture added to the experimental well. The final volume of each well was 10 mL. These plates were incubated for two days at 20 μmol photons m−2 s−1 with a 14 : 10 h light : dark cycle.
To determine cell abundances at the beginning and end of the experiments, 4-mL aliquots were removed from each well and fixed with Lugol’s solution (final concentration = 5%). After a 2-day incubation, the growth rate and bioluminescence of N. scintillans were determined as described in the “Measurement of bioluminescence intensity” and “Measurement of growth rate” sections.
For Experiment 2, dense cultures of P. kofoidii were transferred to seven 250-mL flasks containing A. minutum and filtered seawater. Cells of A. minutum were transferred to seven flasks containing L1 medium without silicate (hereafter L1). After screening the water temperature by pre-incubation, the target temperature was finally established at 5, 10, 15, 20, 25, and 30°C. The procedures for establishment, incubation, and counting in Experiment 2 were the same as those in Experiment 1, except for the target heterotrophic dinoflagellate and its prey species and culture medium (L1 instead of F/2).
For Experiment 3, seven flasks were filled with fresh F/2 medium and A. mediterraneum cells. After screening the effect of temperature by pre-incubation, target temperatures of 10, 15, 20, 25, and 30°C were established. The initial concentrations of A. mediterraneum were achieved by adding a predetermined volume of culture having a known cell abundance to a 38-mL flask and filled with F/2 medium adjusted to each target temperature (final volume = 30 mL). Triplicate culture flasks at each target temperature were established. These flasks were incubated for four days at 100 μmol photons m−2 s−1 and a 14 : 10 h light : dark cycle. At the beginning and end of the experiments, 4-mL aliquots were removed from each flask and fixed with Lugol’s solution. After a 4-day incubation, the bioluminescence and growth rate of A. mediterraneum were determined.
Measurement of bioluminescence intensity
Bioluminescence intensity of each experiment was measured using the method described by Park et al. (2021). In Experiments 1 and 2, after two days of incubation, a 200-μL aliquot of filtered seawater adjusted to each experimental temperature was pipetted into a Corning 96-well white plate (Corning Life Sciences, Amsterdam, Netherlands). Three cells of N. scintillans (or P. kofoidii) under each experimental condition were then isolated from the culture and dispensed into a well containing filtered seawater, with five replicates. Subsequently, a 200-μL aliquot of filtered seawater was placed in a well, with five replicates as control. The 96-well white plate was incubated in darkness for 3 h and at the target temperature conducted for each experiment (Biggley et al. 1969, Krasnow et al. 1980, Lindström et al. 2017). Bioluminescence was chemically stimulated by adding 50 μL of 1 M acetic acid to the well, and stimulated bioluminescence was measured for 200 s using a GloMax Navigator microplate luminometer (Promega, Madison, WI, USA) as described by Park et al. (2021). Experiment 3 involved placing a 200-μL aliquot of a 4-day incubated culture of A. mediterraneum at the target temperature into one well of a 96-well white plate, with five replicates. Bioluminescence intensity was measured following the method described above.
To obtain the bioluminescence intensity per cell per second in each experiment, the mean value obtained from the five control wells was subtracted from the bioluminescence intensity of the cells per second in one well, and then the value was divided by the total number of cells in the well (Park et al. 2021). Using the data on bioluminescence intensity per cell per second in each experiment, the integrated bioluminescence intensity per cell for 200 s were calculated. The mean bioluminescence intensity per cell for 200 s (BLcell; relative light unit [RLU] cell−1) was calculated by averaging the bioluminescence intensity per cell per second for 200 s in five wells. N. scintillans, P. kofoidii, and A. mediterraneum emitted the most bioluminescence in 200 s and thus, BLcell is similar to total luminescence capacity per cell (von Dassow and Latz 2002).
Measurement of growth rate
To determine cell abundances at the beginning and end of the experiments, 4-mL aliquots were removed from each well or flask and fixed with Lugol’s solution (final concentration = 5%). Subsequently, all or >200 cells of a bioluminescent organism and prey were counted in a Sedgewick–Rafter counting chamber.
The specific growth rate of each bioluminescent dinoflagellate, μ (d−1), was calculated as follows:
, where P0 and Pt are the concentrations of each bioluminescent dinoflagellate at 0 and 48 h for N. scintillans and P. kofoidii or at 0 and 96 h for A. mediterraneum, respectively.
Calculation of population bioluminescence intensity of three dinoflagellate species in Korean waters
The population bioluminescence intensities (BLpopul) of N. scintillans, P. kofoidii, and A. mediterraneum were calculated by multiplying their abundance in Korean waters in 1998–2022 by the calculated bioluminescence intensity per cell at the water temperature by linear interpolation (BLcellIT). The abundance data of the three bioluminescent dinoflagellates and water temperature were obtained from Yoo et al. (2002, 2013), Jeong et al. (2011), Lim et al. (2017), Eom et al. (2021), National Institute of Fisheries Science, Korea (2024), and our unpublished data. The values with a cell abundance of N. scintillans equal to or greater than 5,000 cells mL−1 excluded from the analysis.
Statistical analyses
An analysis of variance (ANOVA) was used to test the effects of water temperature on bioluminescence intensity of bioluminescent dinoflagellates. Normality and homogeneity of variance were tested using Shapiro-Wilk’s W and Levene’s tests, respectively. If the data were not normally distributed, data were analyzed using the Kruskal-Wallis test followed by the Mann-Whitney U test with Bonferroni correction (p < 0.05) (Mann and Whitney 1947, Kruskal and Wallis 1952). If variances between treatment groups were not homogeneous, Welch’s one-way ANOVA and the Games-Howell post-hoc test were performed (Welch 1947, Games and Howell 1976). Correlations among the mean BLcell, water temperature, and growth rate were examined using Pearson correlation coefficients or non-parametric Spearman’s rank correlation coefficients. Before applying the correlation tests, the normality of data was confirmed using Shapiro-Wilk’s W test. Statistical analyses were performed using SPSS version 25.0 (IBM-SPSS Inc., Armonk, NY, USA).
RESULTS
Cell shape and growth rates of three species of dinoflagellates at different water temperatures
The cell shapes of N. scintillans NSDJ2010, P. kofoidii PKJH1607, and A. mediterraneum AMYS1807 were affected by water temperatures within the range of 5–35°C. Cells of N. scintillans maintained their normal shape within 5–25°C, but their tentacle and cytoplasmic strands were diminished at 30°C (Fig. 2A–F). Cells of P. kofoidii maintained their normal shape within the range of 5–30°C, but their cytoplasm at 35°C was reduced in volume (Fig. 2G–M). The cytosol of A. mediterraneum became separated from their theca at 5 and 35°C, whereas this species maintained their normal cell shapes at other temperatures (Fig. 2N–T).
Light micrographs of cells of Noctiluca scintillans (A–F) and Polykrikos kofoidii (G–M) incubated for 2 days, and cells of Alexandrium mediterraneum (N–T) incubated for 4 days at the target water temperatures. Scale bars represent: A–F, 100 μm; G–M, 20 μm; N–T, 10 μm.
The specific growth rates of N. scintillans feeding on D. salina at 5 and 10°C were almost zero (−0.01 d−1) and negative (−0.16 d−1), respectively, but increased to 0.54 d−1 at 20°C (Fig. 3A). The specific growth rate of P. kofoidii feeding on A. minutum at 5°C was negative (−0.18 d−1) but rapidly increased to a maximum (0.30 d−1) at 10°C (Fig. 3B). The specific growth rate of A. mediterraneum at 10°C was almost zero (−0.07 d−1) but increased to 0.39 d−1 at 25°C (Fig. 3C).
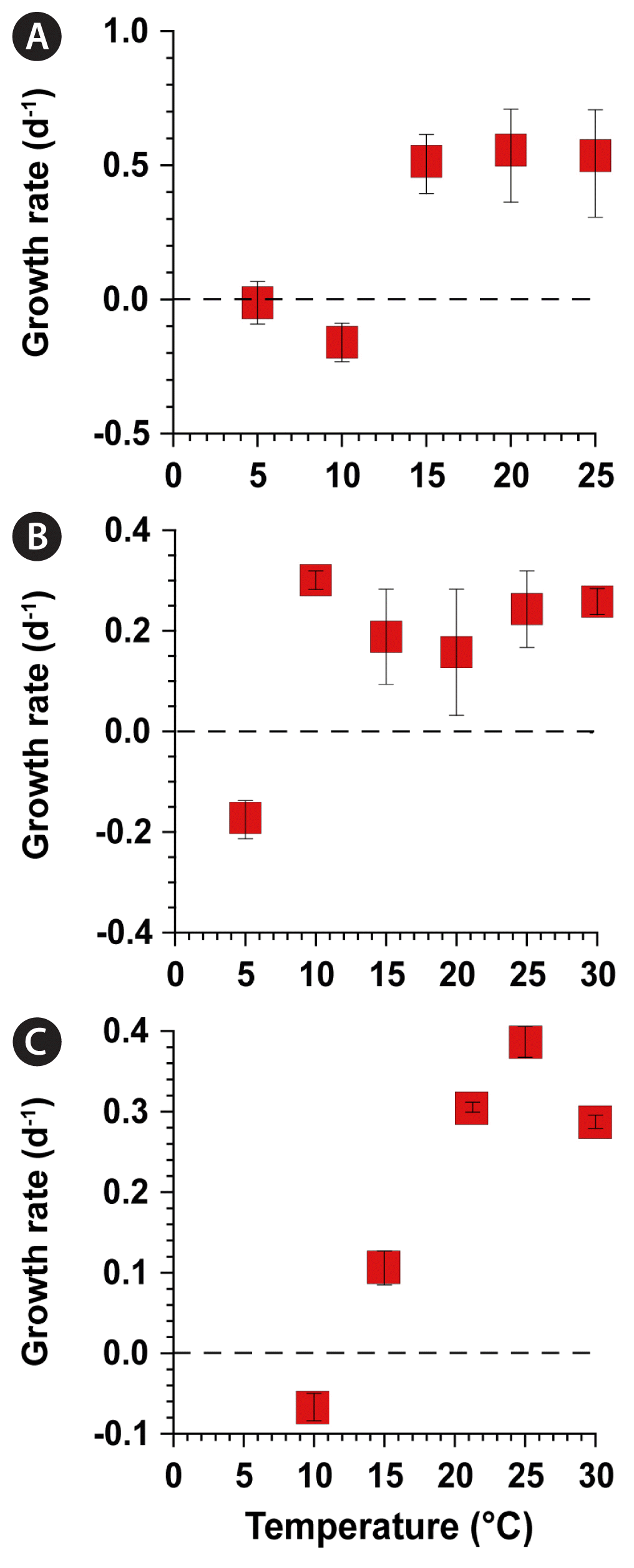
Specific growth rates of Noctiluca scintillans feeding on Dunaliella salina (A), Polykrikos kofoidii feeding on Alexandrium minutum (B), and Alexandrium mediterraneum in F/2 seawater medium without silicate (C) as a function of water temperature. Symbols represent treatment means ± standard error (n = 3).
Effects of water temperature on bioluminescence intensity of Noctiluca scintillans
Mean BLcell of N. scintillans feeding on D. salina was significantly affected by water temperature (Kruskal-Wallis test; H4 = 17.26, p = 0.002). The mean BLcell of N. scintillans was 1.13 × 107–3.91 × 107 RLU cell−1, and the highest mean BLcell was achieved at 5°C (Fig. 4). The values were divided into two subsets (Mann-Whitney U test with Bonferroni correction, p < 0.05). Significant negative correlations were apparent between the mean BLcell of N. scintillans and water temperature (Spearman correlation; rs = −0.82, p < 0.001).
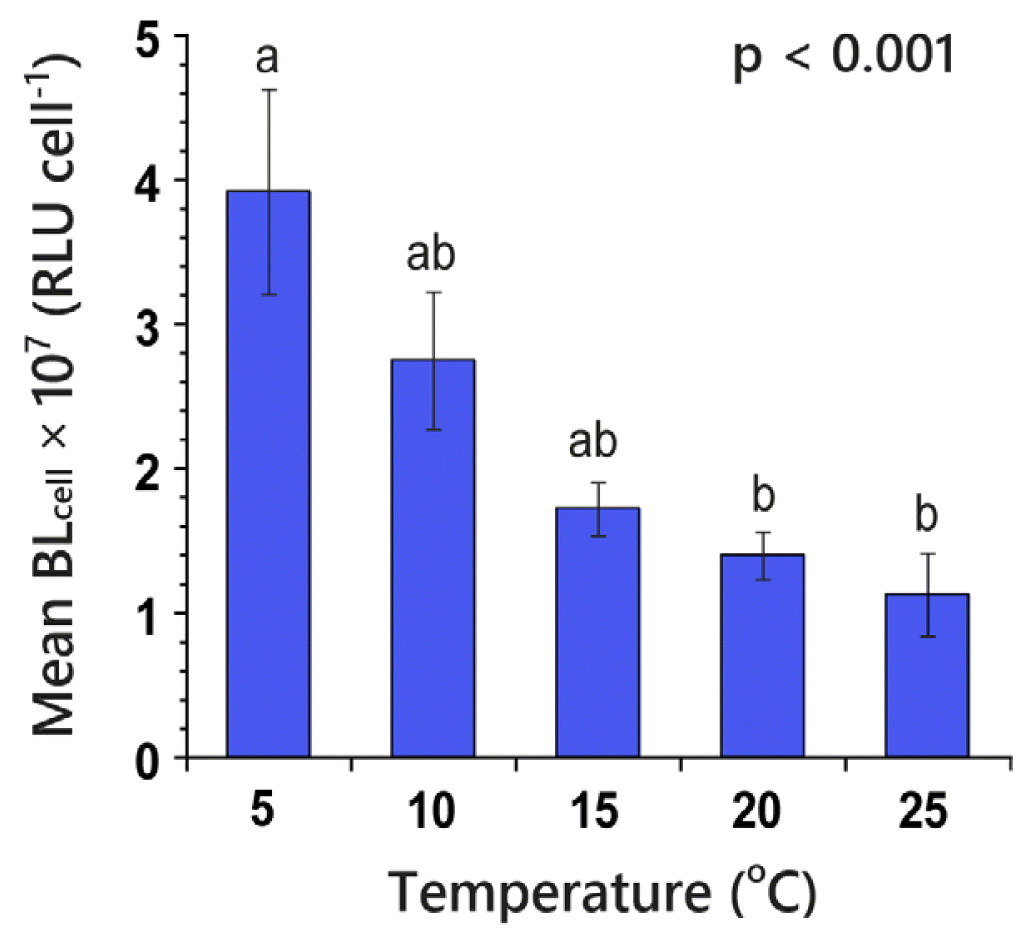
Bar graph of mean 200-second-integrated bioluminescence intensity per cell (BLcell) of Noctiluca scintillans feeding on Dunaliella salina as a function of water temperature ranging from 5 to 25°C. RLU, relative light unit. Mann-Whitney U test with Bonferroni correction after the Kruskal-Wallis test resulted in significantly different subsets for mean BLcell (p < 0.05). Different letters on the top of the bars indicate a significant difference between subsets. Bars represent treatment means ± standard error (n = 5).
Effects of water temperature on bioluminescence intensity of Polykrikos kofoidii
Mean BLcell of P. kofoidii feeding on A. minutum was significantly affected by water temperature (Kruskal-Wallis test, H5 = 19.94, p = 0.001). The mean BLcell of P. kofoidii across the range of 5–30°C was 0.44 × 105–5.19 × 105 RLU cell−1, and the highest mean BLcell was achieved at 15°C (Fig. 5). Bonferroni correction (p < 0.05) revealed that the mean BLcell of P. kofoidii was divided into two subsets.
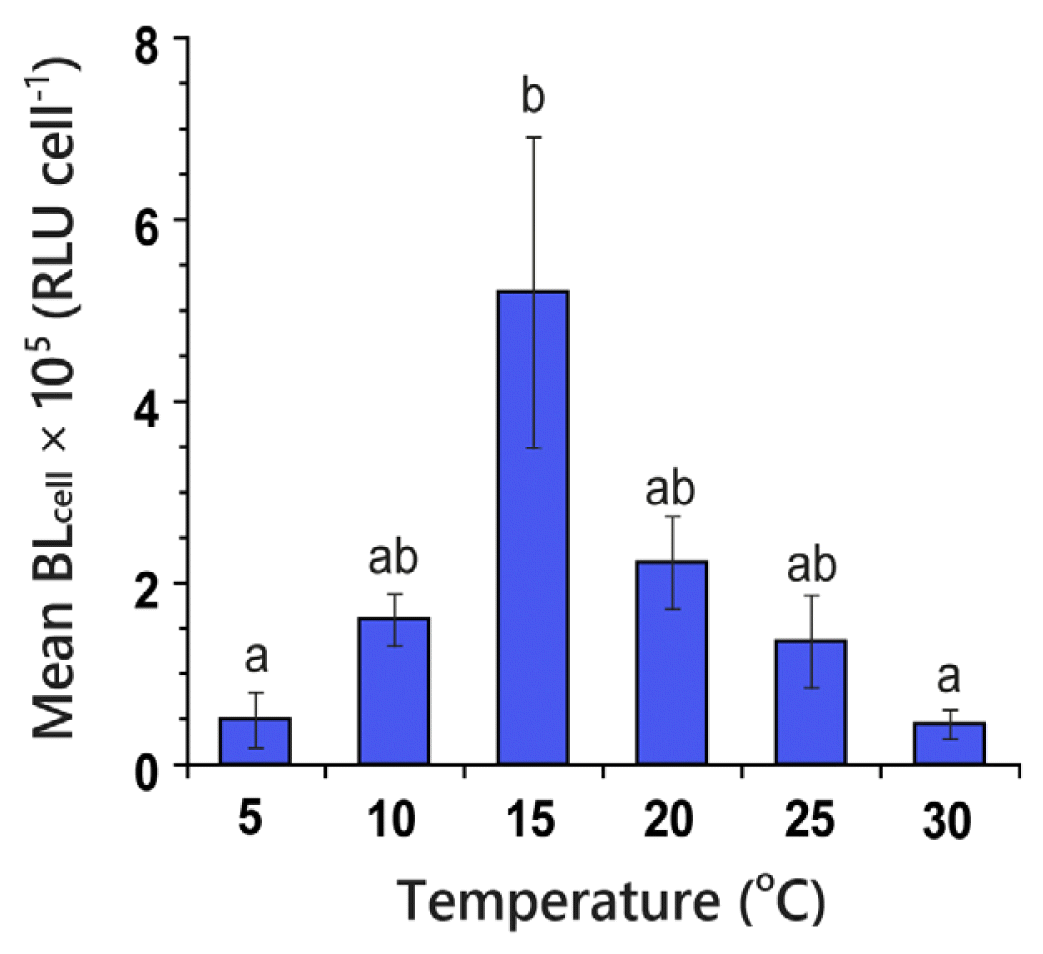
Bar graph of mean 200-second-integrated bioluminescence intensity per cell (BLcell) of Polykrikos kofoidii feeding on Alexandrium minutum as a function of water temperature (°C) ranging from 5 to 30°C. RLU, relative light unit. Mann-Whitney U test with Bonferroni correction after the Kruskal-Wallis test resulted in significantly different subsets for mean BLcell (p < 0.05). Different letters on the top of the bars indicate a significant difference between subsets. Bars represent treatment means ± standard error (n = 5).
Effects of water temperature on bioluminescence intensity of Alexandrium mediterraneum
Mean BLcell of A. mediterraneum was significantly affected by water temperature (Welch’s ANOVA, F4, 9.53 = 24.21, p < 0.001) (Fig. 6). The mean BLcell of A. mediterraneum across the range of 10–30°C was 0.10 × 104–2.20 × 104 RLU cell−1, and the highest mean BLcell was achieved at 20°C. The values were divided into four subsets (Games-Howell post-hoc test, p < 0.05).

Bar graphs of mean 200-second-integrated bioluminescence intensity per cell (BLcell) of Alexandrium mediterraneum as a function of water temperature (°C) ranging from 10 to 30°C. RLU, relative light unit. Games-Howell post-hoc test after the Welch’s ANOVA test resulted in significantly different subsets for mean BLcell (p < 0.05). Different letters on the top of the bars indicate a significant difference between subsets. Bars represent treatment means ± standard error (n = 5).
Correlations between growth rate and bioluminescence intensity of three species of dinoflagellates
Mean BLcell was not significantly correlated with growth of N. scintillans, P. kofoidii, or A. mediterraneum (Pearson correlation; r = −0.85, p = 0.07 for N. scintillans; r = 0.23, p = 0.66 for P. kofoidii; r = 0.40, p = 0.51 for A. mediterraneum) (Fig. 7). The highest mean BLcell of N. scintillans occurred at 5°C, while growth rate was almost zero at that temperature. The highest mean BLcell for P. kofoidii and A. mediterraneum occurred at 15 and 20°C, respectively, where growth rate was the fourth and second-highest, respectively.

Mean 200-second-integrated bioluminescence intensity per cell (BLcell) of Noctiluca scintillans feeding on Dunaliella salina (A), Polykrikos kofoidii feeding on Alexandrium minutum (B), and Alexandrium mediterraneum (C) as a function of growth rate. RLU, relative light unit. Symbols represent the mean and standard error.
Calculation of population bioluminescence intensity of the three dinoflagellates in Korean waters using cell abundance and water temperature
The BLpopul of N. scintillans was 4.71 × 108–4.22 × 1013 RLU L−1 when its abundance was 4.54 × 10–3.50 × 106 cells L−1, and water temperature ranged from 10.9 to 29.3°C in Korean waters between 1998 and 2022 (n = 867) (Fig. 8A). The BLpopul of P. kofoidii was 2.15 × 107–2.28 × 1010 RLU L−1 when its abundance was 1.32 × 102–3.22 × 105 cells L−1, and water temperature ranged from 6.0 to 26.2°C in Korean waters between 1999 and 2021 (n = 146) (Fig. 8B). The BLpopul of A. mediterraneum was 7.26 × 104–7.62 × 107 RLU L−1 when its abundance was 1.25 × 10–1.17 × 104 cells L−1, and water temperature ranged from 10.0 to 29.7°C in 2021 (n = 24) (Fig. 8C).
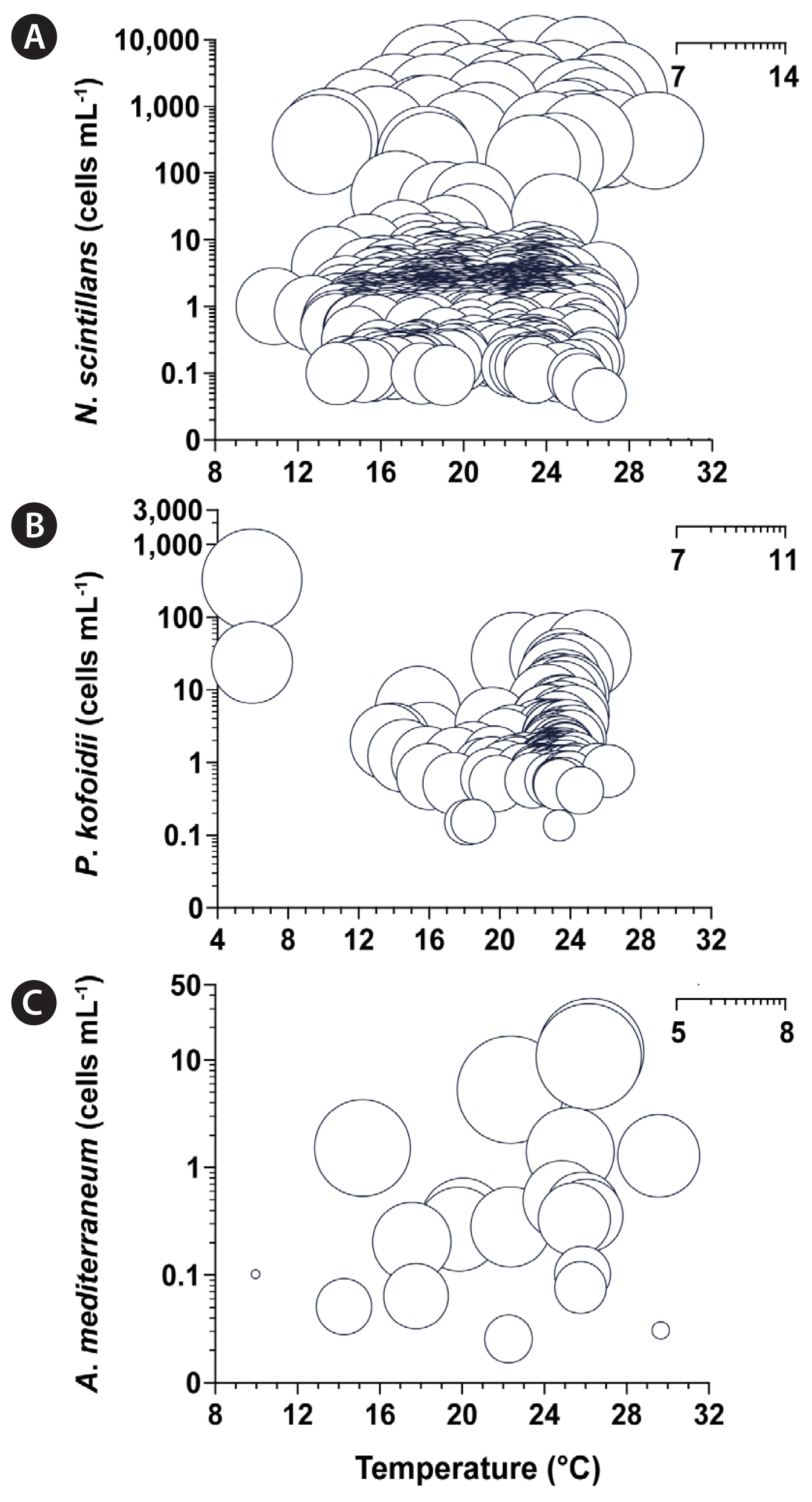
The logarithm of the calculated population bioluminescence intensity (BLpopul) of Noctiluca scintillans (n = 867) (A), Polykrikos kofoidii (n = 146) (B), and Alexandrium mediterraneum (n = 24) (C) using their abundances and water temperatures in Korean waters from 1998 to 2022. The diameter of each circle represents the log (BLpopul) and the center of the circle the cell abundance (y-axis) and water temperature (x-axis) in each sample.
The highest BLpopul of N. scintillans, 4.22 × 1013 RLU L−1, occurred in summer, while that of P. kofoidii, 2.28 × 1010 RLU L−1, occurred in winter (Tables 3 & 4, Fig. 9). The BLpopul of A. mediterraneum was highest, 7.62 × 107 RLU L−1, in summer (Table 5, Fig. 9C). The highest value of BLpopul of N. scintillans was 1,850 times greater than that of P. kofoidii and 554,000 times higher than that of A. mediterraneum.

Estimation of the maximum population bioluminescence intensity of Noctiluca scintillans in three seas of Korea and four seasons

Estimation of the maximum population bioluminescence intensity of Polykrikos kofoidii in three seas of Korea and four seasons
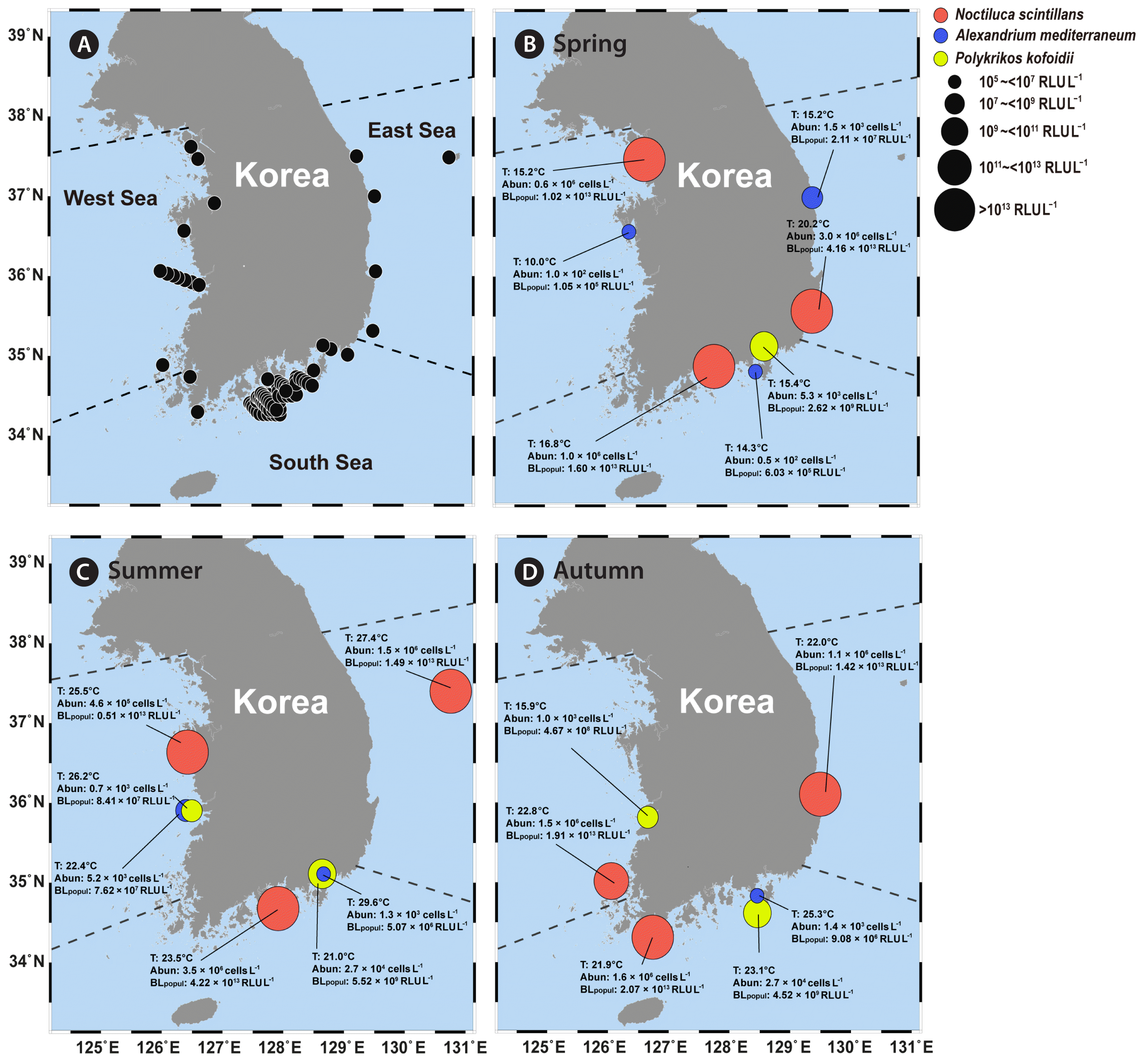
Map of the locations of stations in Korean waters (A) and the maximum population bioluminescence intensity (BLpopul) of Noctiluca scintillans, Polykrikos kofoidii, and Alexandrium mediterraneum in spring (March to May) (B), summer (June to August) (C), and autumn (September to November) (D) in three seas of Korea from 1998 to 2022. Circles indicate the stations at which maximum BLpopul of N. scintillans, P. kofoidii, and A. mediterraneum were obtained. T, temperature (°C); Abun, cell abundance; RLU, relative light unit.

Estimation of the maximum population bioluminescence intensity of Alexandrium mediterraneum in three seas of Korea and four seasons
DISCUSSION
The present study is the first to investigate the effect of temperature on the bioluminescence intensity P. kofoidii and A. mediterraneum. Prior to this study, among the 84 bioluminescent dinoflagellate species, temperature effects on bioluminescence intensity of only five species (N. scintillans, Lingulodinium polyedra, Pyrocystis fusiformis, P. lunula, and P. noctiluca) have been explored (Sweeney 1963, 1981, Kelly 1968, Tett 1971, Jess 1985, Swift et al. 1995, Craig et al. 2003, Cusick and Widder 2020, Park et al. 2021). Thus, the results of the present study extended the number of the bioluminescent dinoflagellate species from 5 to 7 species whose bioluminescence intensity and water temperature effects were explored. Furthermore, the effect of water temperature on the bioluminescence intensities of N. scintillans, L. polyedra, P. fusiformis, and P. noctiluca was investigated at only 10 and 20°C (Sweeney 1981, Jess 1985). The present study examined the effects of temperatures on the bioluminescence intensity of N. scintillans across a broader range of temperatures.
The ratio of the highest BLcell to the lowest BLcell of P. kofoidii in the range of water temperature tested, 11.8, was slightly larger than that of L. polyedra, 10 (Sweeney 1981). Furthermore, the ratio of the highest-to-lowest BLcell of N. scintillans in the range of water temperature tested, 3.5, was similar to that of P. lunula in the temperature range of 10–30°C, 3.0 (Craig et al. 2003). The ratio of the highest-to-lowest BLcell of A. mediterraneum in the range of water temperature tested, 21.0, was considerably larger than that of L. polyedra and P. lunula. Therefore, the effect of temperature on the ratio of the highest-to-lowest BLcell of dinoflagellates is species-specific, and further studies of this ratio across a range of water temperatures are needed.
The growth rates of N. scintillans at 5 and 10°C were negative, but the BLcell of N. scintillans at these water temperatures was higher than that at 15–25°C at which the growth rates were positive. Undivided N. scintillans cells at 5 and 10°C may retain a larger amount of the accumulated luminescent materials such as luciferin and luciferase than divided N. scintillans cells at 15–25°C. However, further study is needed to test this hypothesis.
The BLcell of N. scintillans across the range of 5–30°C in the present study was negatively correlated with water temperature. Chen et al. (2023) reported that the bioluminescence intensity (BLcell) of N. scintillans in the Yellow Sea and Bohai Sea was negatively correlated with temperature across the range of 3–27°C. Therefore, the results of the present study are generally consistent with field observations.
The present study reported, for the first time, an estimation of the bioluminescence intensity of dinoflagellates at seas by using both cell abundance and water temperature. The highest BLpopul of N. scintillans in Korean waters (1998–2021) was 23,000 and 6,886,000 times larger than that of P. kofoidii and A. mediterraneum, respectively. Therefore, N. scintillans populations in Korean waters can produce much brighter bioluminescence than P. kofoidii or A. mediterraneum populations. The highest BLcellIT of N. scintillans was 52 and 1,174 times larger than that of P. kofoidii and A. mediterraneum, respectively. Thus, the ratio of the highest BLpopul of N. scintillans relative to P. kofoidii or A. mediterraneum was much larger than that of the highest BLcellIT of N. scintillans relative to P. kofoidii or A. mediterraneum. The ratio of the maximum abundance of N. scintillans relative to P. kofoidii or A. mediterraneum in Korean waters was 155 or 4,274, respectively (National Institute of Fisheries Science, Korea 2024, our unpublished data). Thus, the combination of the higher BLcellIT and maximum abundance of N. scintillans than that of P. kofoidii and A. mediterraneum resulted in the higher BLpopul of N. scintillans.
The maximum abundance of N. scintillans in Korean waters used in the estimation of its BLpopul was as high as 3.5 × 106 cells L−1. The highest abundance of N. scintillans cultivated in our laboratory was 4.5 × 104 cells L−1 (You et al. 2022). N. scintillans cells tend to float and aggregate at the surface (Tiselius and Kiørboe 1998, Padmakumar et al. 2010). Therefore, during red tides, their abundance in surface waters often becomes extremely high (Mohamed and Mesaad 2007, Padmakumar et al. 2010, National Institute of Fisheries Science, Korea 2024). Patches of red tide of N. scintillans emit a glowing blue light at night, which is easily visible to the naked eye (Padmakumar et al. 2010, Temnykh et al. 2022). In contrast, the maximum abundance of P. kofoidii and A. mediterraneum in Korean waters used in the estimation of its BLpopul was as high as 3.22 × 105 and 1.17 × 104 cells L−1, respectively. Unlike N. scintillans, dense aggregations of P. kofoidii and A. mediterraneum in Korean waters are yet to be reported. N. scintillans caused 125 red tides in the Korean coastal waters from 1981 to 2022 (National Institute of Fisheries Science, Korea 2024). Thus, N. scintillans is likely to be responsible for bright bioluminescence in the surface waters of Korea.
The results of the present study demonstrated that water temperature affects the BLcell, BLcellIT, and BLpopul of N. scintillans, P. kofoidii, and A. mediterraneum. However, the ranges of the abundances of N. scintillans, P. kofoidii, and A. mediterraneum in Korean waters were much wider than those of the BLcellIT in the present study. Therefore, in the estimation of BLpopul of N. scintillans, P. kofoidii, and A. mediterraneum, their abundances in Korean waters may be more important determinants of BLcell, BLcellIT, and BLpopul than water temperature. However, water temperature affects the abundances of N. scintillans, P. kofoidii, and A. mediterraneum and consequently, may affect their BLpopul directly and indirectly.
There are several other bioluminescent dinoflagellates whose abundance is high at seas; the bioluminescent heterotrophic dinoflagellates Protoperidinium spp. and phototrophic dinoflagellates Tripos spp. (previously Ceratium spp.) and Pyrodinium bahamense are sometimes abundant (Lapota and Losee 1984, Lapota et al. 1988, Swift et al. 1995). To understand the bioluminescence produced by dinoflagellate communities, it is worthwhile to investigate the effects of water temperature on their BLcell and estimate their BLpopul in the ocean.
ACKNOWLEDGEMENTS
This research was supported by the National Research Foundation (NRF) funded by the Ministry of Science and ICT (NRF-2021M3I6A1091272; 2021R1A2C1093379; RS-2023-00291696) award to HJJ.
Notes
The authors declare that they have no potential conflicts of interest.