INTRODUCTION
Ocean acidification refers to changes in seawater carbon chemistry that result from the increased influx of anthropogenic CO2 into the oceans (Doney et al. 2009). These changes include decreases in ocean pH, CO32− concentrations, and CaCO3 saturation states, and increases in CO2 and HCO3− concentrations. Given that CO2 and HCO3− are important in supporting algal photosynthesis and carbon metabolism, many autotrophs will likely be impacted by ocean acidification in both pelagic and coastal ecosystems. Though these impacts are not fully understood, photosynthetic organisms that produce calcium carbonate skeletons will generally be negatively impacted by ocean acidification while non-calcifying macroalgae and seagrasses may be affected variously (Kroeker et al. 2013). Understanding species-specific differences in these impacts, especially for near shore benthic macroalgae, may better allow us to predict how coastal ecosystems will be impacted by further changes in ocean acidity.
Macroalgae are dominant components of rocky shores where even though they occupy only a small area of the coastal region, they account for high percentages of coastal carbon cycles due to their high productivity (Duarte and Cebrián 1996, Mcleod et al. 2011, Kim et al. 2015). Most marine macroalgae photosynthesize using C3 pathways and are generally able to overcome limited diffusion / supply and poor affinity of CO2 at the site of CO2 fixation by taking up HCO3− from the seawater to use as a carbon source (Giordano et al. 2005, Raven et al. 2008, 2011). This process is facilitated by carbon concentration mechanisms (CCMs), which are strongly associated with the active transport of HCO3− into the plastid membrane by an enzyme-catalyzed reaction. However, the preferences for different carbon sources that require CCMs are dependent on numerous factors, including the water depth and / or tidal zonation where the macroalgae inhabit (Murru and Sandgren 2004, Hepburn et al. 2011), the total dissolved inorganic carbon (TIC) and / or CO2 concentrations in the water (Giordano and Maberly 1989, Cornwall et al. 2012, Ní Longphuirt et al. 2013), and the species being considered (Maberly 1990, Maberly et al. 1992). Among these factors, increased seawater CO2 may be most important to algal photosynthesis and growth, especially in relation to future ocean conditions.
Most macroalgae that have CCMs rely on the enzyme activity of carbonic anhydrase in order to catalyze the conversion from HCO3− + H+ to CO2 + H2O, which result in photosynthesis being carbon saturated, or nearly so, at the site of carbon fixation under present ocean conditions (Giordano et al. 2005, Falkenberg et al. 2013). This process, however, requires energy for the biophysical transportation of HCO3− into the plasma membrane, it might influence the energy cost within the thallus (Raven et al. 2014). Also, the relative composition of different inorganic carbon species in the coastal waters can variously influence carbon saturation states for macroalgal photosynthesis (Gao et al. 2012, Koch et al. 2013). As a result, the metabolic changes required for photosynthetic acclimation to acidified seawater could similarly vary along with the balance between the energy acquisition and consumption, potentially resulting in negative and / or unpredictable physiological responses within the macroalgae. In contrast, species of macroalgae that rely on only a small contribution of CCM for photosynthesis, do not require the same energy to transport HCO3− into the plasma membrane, but instead rely on ambient CO2 in the seawater and therefore may respond positively to elevated CO2 conditions (Raven et al. 2011, Cornwall et al. 2012, Gao et al. 2012, Koch et al. 2013). These species are regularly exposed to, and thus acclimated for, carbon-limited condition suggesting that inorganic carbon needed for photosynthesis is undersaturated under current ocean conditions, and that exposure to chronic CO2 elevation might result in the enhancement of photosynthesis in them. In this respect, some macroalgae are expected to become more dominant components of their ecosystems under a more acidic ocean while others will decrease in abundance (Hepburn et al. 2011, Johnson et al. 2012). Therefore, understanding the photosynthetic carbon use strategies, combined with knowledge of ocean inorganic carbon saturation states will be integral in predicting ecophysiological and demographic changes in macroalgae under future CO2 conditions (Cornwall et al. 2012).
Many ocean acidification experiments, to date, have relied on tightly controlled stable laboratory conditions and have focused on single species at a time (Widdicombe et al. 2010). Consequently, the results from stable laboratory experiments do not incorporate natural fluctuations in carbon chemistry or other environmental parameters such as temperature, salinity, and irradiance, and therefore may not adequately reflect true ecological responses expected under elevated CO2 conditions in the oceans (Cornwall et al. 2013). Thus, field mesocosm experiments may be required if we are to fully interpret the results of these laboratory experiments and apply them to the proper ecological scales in nature (Riebesell et al. 2010). In this study, experiments were conducted within an outdoor continuous-flowing mesocosm system that maintained natural variations in seawater CO2, temperature, salinity and solar irradiance. The overarching goals of this experiment was to determine how the physiology and ecology of five macroalgae that are dominant components of the Korean coastline are affected by ocean acidification under natural environmental conditions, and to determine the role of external carbonic anhydrase (eCA) in the CCMs of these species. To do this, we traced growth and photosynthetic carbon metabolism in these macroalgae via measurements photosynthesis under ambient and two high CO2 conditions, and ultimately determined how these were linked to eCA activity.
MATERIALS AND METHODS
CO2 manipulation and mesocosm system
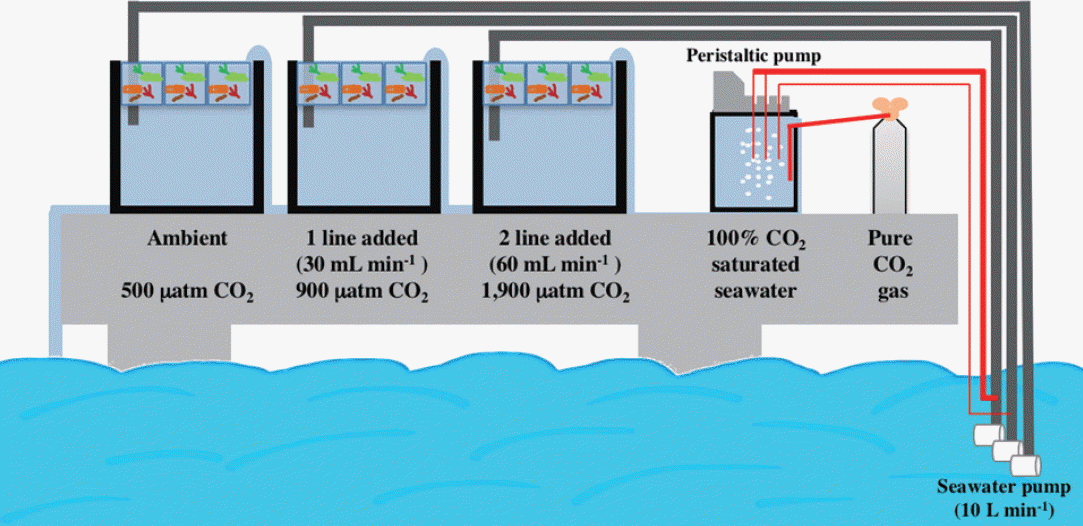
Our mesocosm system simulated ocean acidification by using multi-channel peristaltic pumps to add CO2-saturated seawater into mesocosm inflow pipes, where it then mixed with ambient seawater to create target pH and CO2 conditions (Fig. 1). To do this, CO2-saturated seawater was prepared by bubbling pure CO2 gas into seawater within an 80 L tank, decreasing in seawater pH to approximately 5. A peristaltic pump (Masterflex Pump 7523-57; Cole-Parmer Instrument Co., Niles, IL, USA) was then used to inject this water into the mesocosm inflow pipes at different rates to create two elevated CO2 conditions, with injection rates established by varying the number of injection lines associated with each mesocosm. Ambient seawater (ca. 500 μatm CO2) was pumped at a rate of 10 L min−1 into the mesocosms using 12-V submersible bilge pumps (capacity: 800 GPH/3,028 LPH; Rule Industries Inc., Burlington, MA, USA), and injection rates of CO2-saturated seawater were 30 mL min−1 and 60 mL min−1 for mesocosms. The seawater CO2 concentrations were roughly raised to 2× ambient (resulting in ca. 0.2 pH unit decrease) and 4× ambient (resulting in ca. 0.5 pH unit decrease), respectively (Table 1). Other environmental parameters varied naturally under local conditions. These included natural fluctuations in baseline seawater CO2 (484–518 μatm CO2), temperature (20–23°C), salinity (30–32 psu), and daytime integrated irradiance (13.3–25.1 mol photons m−2 d−1). In situ nutrient levels in the seawater that was pumped into the mesocosms were 3.09 ± 0.91 μM (nitrite + nitrate), 0.62 ± 0.30 μM (phosphate), and 13.95 ± 5.54 μM (silicate) (data provided by Dr. Jang PG).
The seawater pH (National Bureau of Standards [NBS] scale) within the mesocosms was measured at least five times during each day of the experiment using a pH meter (PHM 210; Radiometer, Copenhagen, Denmark) that was calibrated using the National Institute of Standards and Technology (NIST) standard reference material. The differences in pH among the CO2 treatments were maintained over the two-week experiment [ambient (control) = 7.95 ± 0.03; 2× ambient (1 line added) = 7.72 ± 0.06; 4× ambient (2 lines added) = 7.45 ± 0.15]. In addition, seawater carbon chemistry was measured using potentiometric acid titration as described by Millero et al. (1993). TIC and AT (total alkalinity) were determined using the methods described in Hernández-Ayón et al. (1999) and Millero et al. (1993), respectively. The CO2sys basic software then used two parameters of the carbonate system (AT and TIC in this study) to calculate pCO2 (partial pressure of CO2), HCO3−, and CO32− in the seawater (Lewis and Wallace 1998). The precisions of our TIC and AT estimate were checked with CRMs (certified by A. Dickson, Scripps Institution of Oceanography, San Diego, CA, USA), and were approximately ±5 μmol kg−1 and ±2 μmol kg−1, respectively (Kim et al. 2013b).
Sample collection
Five locally abundant species of macroalgae (namely the green algae Ulva pertusa and Codium fragile, the brown algae Sargassum horneri and S. thunbergii, and the red alga Prionitis cornea) were selected for our experiments. These species are widely distributed on Korean rocky shores, with the relative abundances of S. thunbergii, C. fragile, and P. cornea varying from upper to lower tidal zones. In contrast, U. petusa occurs irrelevant to tidal height (Choi and Kim 2004), while S. horneri is commonly distributed subtidal areas where it forms in a forest-like brown alga. Algal samples were collected from 2–5 m water depth near Jangmok on the southern coast of Korea (34.6° N, 128.5° E; South Sea Institute of the KIOST) on Sep 30, 2010, and transported into the mesocosm system described above, which was located adjacent to the sampling site. The macroalgae were immediately placed in approximately 0.5 ton open top outdoor mesocosm tanks (length 120 cm × width 85 cm × height 48 cm) equipped with continuous-flowing ambient seawater for three days to allow the algae to acclimate to the mesocosm environment. Following this, 15 individuals of each species (approximately 0.6 g fresh weight of U. pertusa; 2.5 g of C. fragile; 1.0 g of S. horneri; 2.0 g of S. thunbergii; 0.5 g of P. cornea) were transferred to mesh plastic cages (length 44 cm × width 32 cm × height 20 cm), and three cages were placed into each experimental mesocosm containing the three different CO2 conditions (ambient, 2× ambient, 4× ambient). Three plastic cages (n = 3) were randomly placed within each mesocosm tank and their positions within the tanks rotated every day. The plastic cages were fixed at the seawater surface and the algae within the cages were mixed frequently to avoid prolonged shading and heterogeneity of water chemistry. The algae were then held within the mesocosms for two weeks in order to examine the impacts of elevated CO2 on their photosynthesis. All samples chosen for analysis were selected randomly from the three plastic cages and measurements were performed in triplicate.
Photosynthesis (chlorophyll a fluorescence and net O2 production)
Samples were exposed to natural fluctuations in light over the two-week experiment and chlorophyll a fluorescence was measured 20 times on each of three individuals at each treatment between dawn and dusk (06:00–20:00 KST), a period that included a wide range of light intensities (0–1,600 μmol photons m−2 s−1). Effective quantum yield of PSII (ΦPSII) was measured by in vivo chlorophyll a fluorescence using a Diving-PAM (Walz, Effeltrich, Germany). The ΦPSII was measured after exposure to in situ irradiance condition. A leaf distance clip was equipped on the fiber optic to maintain the light exposed area and distance from the samples surface. The ΦPSII was calculated as ΦPSII = ΔF/Fm′ = (Fm′ − F) / Fm′, where F and Fm′ represent the steady-state fluorescence and maximum fluorescence measured in the light, respectively. All ΦPSII measurements were obtained by exposing the samples to a saturation pulse of light followed by different intensities of natural irradiance, and those results represent the apparent efficiency of open PSII reaction centers (Kim et al. 2013a). Relative electron transport rates (rETRs) were calculated as rETR = ΦPSII × irradiance, and steady-state light response curves (LCs) were constructed as rETR-I curves.
In situ incubation experiments were conducted from dawn to dusk (06:00–20:00 KST) in order to determine steady-state photosynthetic O2 evolution and consumption rates under natural irradiances, which were measured using planar oxygen sensor spots (SP-PSt3) and a FIBOX 3 system (PreSens; GmbH, Regensburg, Germany). Working with one individual from each of the five species and three CO2 treatments (i.e., 15 incubation flasks) at a time, thalli of each macroalga (0.2 g fresh weight of U. pertusa, 1.0 g of C. fragile, 0.3 g of S. horneri, 0.5 g of S. thunbergii, and 0.1 g of P. cornea) were put into separate 80 mL Corning cell-culture flasks and oxygen concentration was recorded every 30 min under natural light conditions, which was determined to be sufficient time for detecting production and / or consumption of oxygen during photosynthesis and / or respiration. Following this, the algal samples were removed from the flasks, the water replaced, and new 30-min incubations done. Because this resulted in ~30-min time gaps between replicate measurements for each species-CO2 combination and all photosynthetic measurements were conducted under ambient light conditions, which varied naturally between sample runs, the data for the three macroalgae from each species-CO2 combination could not be considered replicates and thus photosynthetic parameters could not be represented by their means and standard deviations. Rather, photosynthetic data for the three samples from each treatment combination were represented by independent light response curves, each obtained under a range of irradiances (0–1,500 μmol photons m−2 s−1), and photosynthetic parameters were calculated for each of the independent P-I (net photosynthesis-irradiance) curves. The three light responses curves for each species-CO2 combination are represented in electronic supplementary materials (Supplementary Figs S1 & S2). Solar irradiance was recorded using a LI-190 2π PAR sensor connected to a data logger (LI-1400; LI-COR, Lincoln, NE, USA) during the PAM and net photosynthesis measurements and was used to construct the light responses curves, with the data standardized by sample fresh weight (g).
To identify photosynthetic traits, LCs and P-I curves were fitted to a double exponential decay function with a non-linear regression algorithm (Platt et al. 1980). Photosynthetic parameters of LCs (rETRm,LC, maximum relative electron transport rate; α,LC, electron transport efficiency; and Ek,LC, light-saturation coefficient of LCs) and P-I curves (Pmax, maximum net photosynthesis rate; α, photosynthetic efficiency; and Ek, irradiance at the onset of light saturation) were determined using the least squares curve fitting technique included with the software Grapher ver. 9.6 (Golden Software Inc., Golden, CO, USA).
Growth rate
The growth rates of the macroalgae were estimated by measuring changes in the fresh wet weights of three replicate fragments of each species after two weeks in the mesocosms; beginning fresh weights were 0.2 g for U. pertusa, 1.0 g for C. fragile, 0.3 g for S. horneri, 0.1 g for S. thunbergii, and 0.3 g for P. cornea. The specific growth rate (SGR) of each alga was calculated as: SGR (d−1) = ln (WT / W0) / (DT − D0), where WT and W0 represent the sample fresh weights at DT (after 2 weeks) and D0 (the initial day), respectively.
Inhibition of eCA
Inhibition of eCA was estimated by gross oxygen production in the presence and absence of the eCA enzyme inhibitor acetazolamide (60 μM of AZ; Sigma-Aldrich, St. Louis, MO, USA) (Israel and Hophy 2002, Kang et al. 2016). To do this, a stock solution of 40 mM AZ was prepared and then diluted 800-fold by adding it to filtered seawater within a ~80 mL water-jacketed respiration chamber. Light was consistently provided to the chambers with intensity of 200 μmol photons m−2 s−1 by a halogen lamp (KL2500LCD; Schott, Elmsford, NY, USA), and the temperature within the chamber held constant at 22°C. The seawater within the chamber was mixed using a magnetic stirring bar to prevent boundary layer formation. The chamber was then used to measure oxygen production and consumption rates (gross oxygen production) of three replicate samples of each algal species (Kim et al. 2011). Oxygen production was measured using a 2 mm oxygen-dipping probe (DP-PSt3) with a coated foil sensor that was connected to a precise fiber optic oxygen transmitter (FIBOX 3 Oxygen Meter; PreSens GmbH, Regensburg, Germany), and the oxygen changes within the chamber was continuously monitored by personal computer for 20 min under dark and light conditions before and after adding AZ. The eCA inhibition rate of gross photosynthesis (i.e., % of reduced gross photosynthesis by the addition of AZ) was then estimated for each species under each CO2 treatment.
Statistical analysis
All statistical analyses were performed using SPSS ver. 21 (IBM Corp., Armonk, NY, USA). All data met assumptions of normality and equal variances, as determined by Shapiro-Wilk normality and Levene’s homogeneity of variance tests, respectively, except for inhibition rate of eCA which used AZ. In cases where homogeneity of variances was rejected (inhibition rate of eCA), we used Welch analyses of variance (ANOVA) test to check for consistency among the treatments. Photosynthetic parameters of the LCs and P-I curves, inhibition of eCA activity and growth rates of the macroalgae were each compared among the three CO2 conditions and five species using separate two-way Model I ANOVAs. Following this, Tukey’s post hoc multiple comparisons were used to identify specific difference in photosynthetic P-I curve and LCs parameters, and growth among different levels of CO2 concentrations when the ANOVAs identified those factors to be significant (p < 0.05). Paired t tests were used to test for differences in photosynthesis before versus after adding acetazolamide to the incubation chamber.
RESULTS
Chlorophyll a fluorescence and net photosynthetic rate
The five macroalgal species examined in our study exhibited different pattern of rETR-I curves (LCs), but were not noticeably impacted by elevated CO2 concentrations (Table 2, Supplementary Fig. S1). Further, the LCs did not show inhibition (down-regulation) of photosystem II for any of the macroalgal species under high light intensities. C. fragile and P. cornea, which exhibited the highest rETRm,LC and Ek,LC values, did not reach light saturation under our experimental irradiances. In contrast, U. pertusa exhibited the lowest levels of rETRm,LC (195.47 ± 49.28) under ambient conditions. The two Sargassium species showed relatively higher α,LC than other species (0.945 ± 0.117 for S. thunbergii and 0.809 ± 0.059 for S. horneri), but there were no significant differences among the other species (Tukey’s: p > 0.05). The light-saturation coefficients determined by the LCs varied among the macroalgal species under in situ light intensities. This indicates that apparent photochemical activity was not impacted by high CO2 conditions under natural irradiances. Together, photosynthetic parameters of LCs were not significantly impacted by CO2 and / or macroalgal species except for α,LC which varied among the species (F4,30 = 9.426, p < 0.001) (Table 3).
In contrast to LCs, net photosynthetic rates varied slightly among both the macroalgal species and three CO2 conditions (Table 2, Supplementary Fig. S2). Specifically, Pmax varied significantly among the macroalgal species (F4,30 = 48.862, p < 0.001) and interacted with elevated CO2 (F8,30 = 2.526, p = 0.031), but did not vary among the CO2 treatment itself (F2,30 = 0.507, p > 0.05) (Table 3). There was no significant individual or combination effects of CO2 and macroalgal species on the photosynthetic parameters of α, and Ek except for macroalgal species for α (F4,30 = 2.768, p = 0.045) and CO2 for Ek (F2,30 = 3.530, p = 0.042), respectively. Specifically, under ambient conditions, P. cornea exhibited the highest Pmax (512.72 ± 103.77 μmol O2 g−1 FW h−1), while C. fragile exhibited the lowest Pmax (52.79 ± 18.40 μmol O2 g−1 FW h−1). Under elevated CO2, U. pertusa and S. thunbergii exhibited obvious enhancement of Pmax compared to ambient conditions. Pmax in U. pertusa was 21% and 37% and in S. thunbergii was 15% and 51% higher under 2× and 4× ambient CO2 conditions relative to ambient condition, respectively. In contrast, most noticeable reduced photosynthesis occurred in P. cornea (reduced 3% and 30% for Pmax and 29% and 24% for α under 2× and 4× ambient CO2 conditions relative to ambient condition, respectively). Photosynthesis in S. horneri did not vary substantially among the CO2 levels, and C. fragile exhibited the lowest overall photosynthetic capacity compared with the other species, together making it difficult to distinguish between experimental treatments.
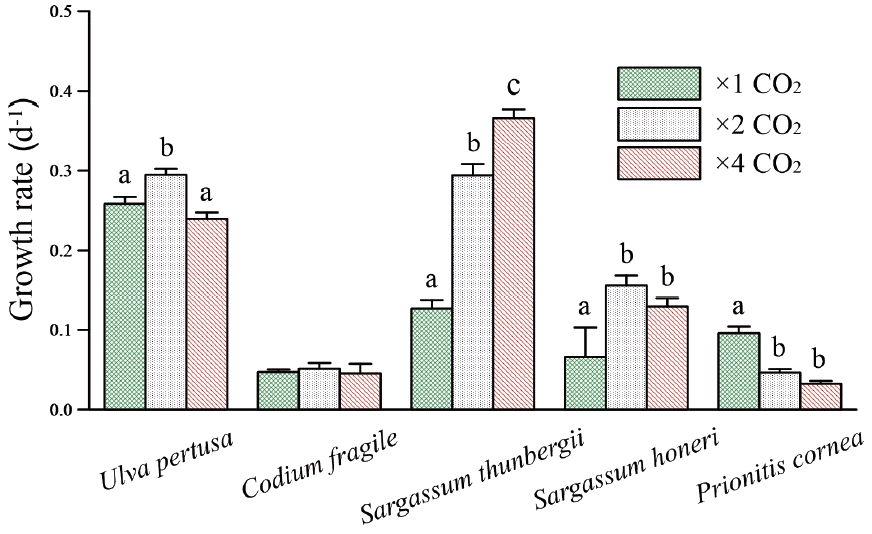
Growth rate
Growth rates varied significantly among the five macroalgal species (F4,30 = 600.846, p < 0.001) and three CO2 treatments (F2,30 = 65.986, p < 0.001) (Table 4, Fig. 2). However, the impact of elevated CO2 on growth was not consistent among the algal species (Species × CO2: F8,30 = 68.211, p < 0.001), indicating that the different species respond differently to elevated CO2. Specifically, growth in the two species of green algae responded very differently to elevate CO2, with U. pertusa exhibiting the highest growth rates under ambient CO2 condition and C. fragile exhibiting the lowest growth rates. U. pertusa growth increased significantly under 2× ambient relative to ambient condition (Tukey’s: p < 0.01) but not under 4× ambient condition (Tukey’s: p > 0.05), while growth in C. fragile remained unaffected by elevated CO2 (Tukey’s: p > 0.05). Growth in U. pertusa increased with elevated CO2 level (with the maximum observed under 2× ambient conditions), which growth in C. fragile remained unaffected by changes in CO2. Furthermore, growth in both S. thunbergii and S. honeri increased significantly under 2× ambient CO2 compared with ambient conditions (Tukey’s: p < 0.001 and p < 0.01, respectively). In contrast, growth in S. thunbergii continued to increase under 4× ambient conditions (Tukey’s: p > 0.01) while growth in S. honeri remained unchanged relative to 2× ambient conditions (Tukeys: p = 0.476). Growth rate in P. cornea decreased significantly under both 2× and 4× ambient relative to ambient condition (Tukey’s: p > 0.001), but it did not differ between the two elevated CO2 treatments (Tukey’s: p = 0.054).
Inhibition of eCA
All species exhibited significant reductions in gross photosynthesis after adding the eCA inhibitor (Fig. 3), but differed in the strength of eCA inhibited photosynthesis. Overall, inhibition of eCA activities varied significantly among the five macroalgal species (F4,30 = 18.911, p < 0.001) but not among the three CO2 concentrations (F2,30 = 1.941, p > 0.05) (Table 5). The relative impact of increased CO2 concentrations on eCA inhibition also varied significantly among the five macroalgal species (F8,30 = 2.457, p < 0.05). Specifically, U. pertusa, S. horneri, and P. cornea exhibited relatively high inhibition of gross photosynthesis compared with C. fragile and S. thunbergii when examined under ambient CO2 conditions, but these results were varied under elevated CO2 conditions. The photosynthetic rates of U. pertusa and P. cornea were 490 and 433 μmol O2 g−1 FW h−1 when examined under ambient conditions, but decreased 73% and 81%, respectively, after eCA was inhibited (Fig. 3). Although they were not significantly different, eCA inhibition rates in U. pertusa increased by 79% and 96% when examined under 2× and 4× ambient conditions, respectively. S. horneri exhibited strong eCA inhibition, with a 95% decrease in its photosynthetic rate following the addition of eCA under ambient conditions, and a 56% and 72% decrease when examined under 2× and 4× ambient conditions, respectively. C. fragile and S. thunbergii exhibited roughly a 46% decrease in photosynthesis following the addition of CA, and a lower depression of eCA activity compared to other species under ambient condition. Further, eCA inhibition in S. thunbergii was significantly decreased when exposed to high levels of CO2 relative to ambient condition (Tukey’s: p < 0.05). In contrast, eCA inhibition in C. fragile, U. pertusa, and P. cornea did not change under elevated CO2 conditions (Tukey’s: p > 0.05).
DISCUSSION
Field-based mesocosms have been recognized as ideal tools for evaluating ecological responses by organisms to changes in their environment (Petersen et al. 2009). In contrast, mesocosm systems for studying the impacts of ocean acidification have proven difficult to maintain CO2 conditions at desired levels while allowing other environmental factors to fluctuate naturally (Havenhand et al. 2010, Widdicombe et al. 2010). For example, several field mesocosm studies have manipulated CO2 concentrations by bubbling specific CO2-air gas mixtures into seawater within enclosed mesocosm systems (e.g., Alexandre et al. 2012, Olabarria et al. 2013), but these studies tend to hold CO2 concentrations constant at these levels and thus do not allow natural daily and / or diurnal variability in CO2 or other environmental variables. To address this problem, our mesocosm system relies on continuous-flowing seawater with a turnover rate within each tank of less than 45 min, and incorporates natural fluctuations in seawater temperature, salinity, nutrient supply, oxygen and carbon chemistries, and ambient irradiance. We believe this design may be more powerful for studying how macroalgae will respond to future CO2 levels in the complex coastal environment. However, a limit of this mesocosm study is the small number of mesocosm tanks (replicates) for each CO2 treatment. First, it is logistically difficult to build and maintain these tanks, and we lack manpower to measure each additional (replicates) tanks. Such problems with pseudoreplication (sensu Hurlbert 1984) have long been recognized as problematic when interpreting the results of ecological studies, and have been particularly prevalent in mesocosm studies where the number of tanks and / or available space is often limited. However, given the efficiency of this system in manipulating CO2, we believe our mesocosm design is ideal for evaluating impacts of ocean acidification on macroalgal photosynthesis (Havenhand et al. 2010, Widdicombe et al. 2010), but we caution the reader to recognize the lack of replication of mesocosm tanks when applying these results to the greater coastal zone.
Chlorophyll a fluorescences were measured under seminatural conditions, which incorporated daily irradiance cycles. Our results suggest that PSII photochemical performance is not necessarily influenced by variability in CO2 concentration under conditions of fluctuating light intensities in nature, and it might be hard to detect changes in photochemical activity using seminatural mesocosm studies with acidified seawater. All photo-physiological characteristics were associated with specific times and irradiances, and various responses of photo-physiological changes have been reported for the marine autotrophs (Kim et al. 2013a). For example, changes of chlorophyll a fluorescence have been observed under well-controlled laboratory experiments (e.g., pelagic organisms: Fu et al. 2007, Sobrino et al. 2008; and benthic plants: Xu and Gao 2012, Oilschläger and Wiencke 2013) or in mesocosm studies (e.g., Connell and Russell 2010, Olabarria et al. 2013). Results from these studies indicate that some benthic plants do not respond strongly to elevated CO2 conditions (e.g., Alexandre et al. 2012, Hofmann et al. 2012a, 2012b). However, there are very few studies are aware of where this has been examined under natural outdoor irradiances.
Although chlorophyll a fluorescence was not affected by ocean acidification conditions, macroalgal photosynthesis and growth did vary in response to elevated CO2. Specifically, U. pertusa and S. thunbergii exhibited increased photosynthetic rates under high CO2, suggesting that the ambient carbon pool is limited (undersaturated) for photosynthesis. These results may be closely connected with improving photosynthesis in relation to energy allocation (Kim et al. 2013a). From the comparing two of our photosynthesis results (chl a fluorescence and net photosynthesis), the energy allocation for photosynthesis between photochemical properties and O2 production is disproportionate under high CO2 conditions, a tendency that was already established for pelagic autotrophic organisms (Sobrino et al. 2008, Kim et al. 2013a). In our study, energy utilization efficiency for the photosynthesis was obviously enhanced under high CO2 in U. pertusa and S. thunbergii. Additional parameters to determine photosynthetic rates using electron transport rates, which include photorespiration, Mehler reactions and nitrate assimilation, are complicated and highly affected by high CO2 conditions (Baker and Oxborough 2004). However, C. fragile did not respond to changes in CO2 concentration with respect to photosynthesis (photochemical activity and O2 production) and growth, because these species are presumably already carbon saturated under ambient conditions. Consequently, TIC saturation states for photosynthesis in the seawater largely influence the growth of some macroalgal species, but not others. In contrast, P. cornea showed decreased net photosynthesis under the highest CO2 conditions, and it might be that this species is very sensitive to decreases in pH.
Our eCA inhibition results under ambient CO2 suggest different species of macroalgae have different strategies of carbon acquisition for eCA. In this study, two groups of macroalgae were roughly identified based on their eCA inhibition results under ambient CO2 conditions; one that describes a highly eCA depressed group (including U. pertusa, S. horneri, and P. cornea), and the other that describes a less eCA sensitive group (including C. fragile and S. thunbergii). These properties of carbon acquisition under ambient condition are well known from previous physiological studies (e.g., Maberly et al. 1992, Koch et al. 2013) and are in agreement with our results. Also these characteristics could be used to predict how these macroalgal species might acclimate physiologically to elevated CO2 environments. For example, S. thunbergii and S. honeri exhibited significantly reduced inhibition of eCA under high CO2 conditions even though these two species have different carbon acquisition properties under ambient condition, suggesting they could take an advantage of elevated CO2 by saving energy through the depression of eCA. Consequently, growth rates in the two species of brown algae increased under high CO2 condition compared to ambient condition. In contrast, three other species examined here, U. pertusa, C. fragile, and P. cornea, did not take advantage of elevated CO2 with respect to energy cost for eCA modulation. The eCA inhibition rate of U. pertusa and P. cornea is very high regardless of CO2 concentration, so their energetic cost remained same or higher than ambient under high CO2 condition. Growth of U. pertusa was not increased under highest CO2 concentration (4× ambient) even though photosynthesis was enhanced compared to ambient condition owing to energy cost for eCA modulation. All data on photosynthetic and eCA metabolisms seem to be connected to the growth dynamics of marine macroalgae. P. cornea was the only exception that did not show changes in eCA inhibition rates, and its photosynthetic rate and growth decreased under the elevated CO2 conditions. We believe this is most likely due to low pH stress as has been previously observed in other red algae such as Porphyra linearis (Israel et al. 1999).
From an ecological perspective, the five macroalgal species have different carbon acquisition strategies that seem to vary along depth and zonation gradients (Hepburn et al. 2011). S. thunbergii is commonly distributed in the upper subtidal region of our study area where it occasionally experiences exposure to the air where only CO2 is available as a carbon source (Kim et al. 1998). Forest-like brown algae such as S. horneri also experience air exposure that can be unrelated to changes in tidal levels (Golléty et al. 2008), and these species may gain an advantage in their competition with turf algae under high CO2 conditions (Olabarria et al. 2013). The highest photosynthetic rates among the five species were observed in S. thunbergii and were correlated with high tolerance to desiccation during exposure to the atmosphere, with a fast diffusion of atmospheric CO2 (Ji and Tanaka 2002). Two brown algal species (S. horneri and S. thunbergii) are frequently exposed to atmospheric CO2, therefore photosynthetic activity could be stimulated by increased CO2 through passive CO2 transport. This tendency is supported by previous studies (e.g., Brown et al. 2014) that report growth of the forest-forming kelp species, Macrocystis pyrifera, is significantly stimulated by elevated CO2 under mesocosm conditions. This also supports the findings of Ní Longphuirt et al. (2013) who demonstrated that some brown algae show significant photosynthetic enhancement when atmospheric CO2 is increased by a natural occurring CO2 vent. Specifically, the abundance of brown algae increased significantly near the natural CO2 vents system, which is also in agreement with our results (Hall-spencer et al. 2008, Johnson et al. 2012).
In contrast to the brown algae, the two green algal species showed little to no responses to the increased CO2 conditions within our mesocosms. Large amounts of physiological information on the Ulva thallus under elevated CO2 conditions were available from previous studies (e.g., Kang et al. 2016), but there is no comparable result for C. fragile. It is generally known that growth of ulvoid species is stimulated under high CO2 and sufficient N conditions (Gordillo et al. 2001, 2003), but dissolved inorganic nitrogen concentrations were relatively lower than the nitrogen conditions used in previous physiological researches. Obviously, growth of U. pertusa was stimulated less than brown algae, and C. fragile did not alter their growth under high CO2 conditions. These two genus act as opportunistic and / or invasive species in coastal areas of temperate region, thus they play an important role in dynamics of macroalgal community fluctuations (Kang et al. 2014, Kang and Kim 2016). If our results represent expressive responses of macroalgae to ocean acidification, blooms of two green algal species could be masked and / or depressed due to over-stimulation of brown algal growth.
In summary, this outdoor flowing-through mesocosm study was conducted to identify the effects of elevated CO2 on the photosynthetic activities (PSII photochemical activity and O2 production) and growth metabolisms in marine macroalgae. While previous studies have suggested positive impacts of ocean acidification on metabolic changes in macroalgae, data describing the comparison between photo-physiology and growth metabolism have not been comparatively sufficient. Thus, we investigated physiological responses of five species of macroalgae under different ocean acidification conditions, and our results represent the tracing of this energetic metabolism under high CO2 conditions. This includes tracing photo-physiological changes associated with the harvesting of light energy to the growth metabolism required for building macroalgal vegetation. Our key finding is that species-specific photosynthetic inorganic saturation states and eCA inhibition are closely related to growth in responses to high CO2 environments, and these results could be crucial in predicting ecophysiological responses of temperate marine macroalgae in future ocean conditions. In this respect, some macroalgal species can be more positive to CO2 enhancement than others, resulting in increase of photosynthesis and growth under elevated CO2 conditions. Based on our results, we suggest that, productivity of S. thunbergii might exceed that of U. pertusa, resulting in S. thunbergii becoming competitively dominant species in this temperate benthic community in the future coastal ocean.
SUPPLEMENTARY MATERIAL
Supplementary Fig. S1. Steady-state light response curves (LCs) of Ulva pertusa, Codium fragile, Sargassum thunbergii, Sargassum horneri, and Prionitis cornea calculated by effective quantum yield of PSII and in situ irradiance under the ambient and two ocean acidification conditions (www.e-algae.org).
Supplementary Fig. S2. Net photosynthesis vs. irradiance (P-I) curves of Ulva pertusa, Codium fragile, Sargassum thunbergii, Sargassum horneri, and Prionitis cornea under the ambient and two ocean acidification conditions (www.e-algae.org).